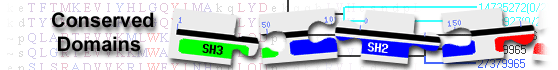oxygenase domain of ten-eleven translocation (TET)2 methylcytosine dioxygenase and similar ...
3-447
0e+00
oxygenase domain of ten-eleven translocation (TET)2 methylcytosine dioxygenase and similar proteins; TET2 is involved in DNA demethylation through iteratively oxidizing 5-methylcytosine (5mC) into 5-hydroxymethylcytosine (5hmC), 5-formylcytosine (5fC) and 5-carboxylcytosine (5caC). Human TET2 (and TET1) have been shown to be more active on 5mC-DNA than 5hmC/5fC-DNA substrates. TET proteins contain a C-terminal catalytic domain which consists of a cysteine-rich region and a double-stranded beta-helix (DSBH) fold. TET2 acts as a tumor suppressor in hematopoiesis; mutations/deletions of the TET2 gene frequently occur in multiple spectra of myeloid malignancies. TET2 (and TET3) promoters are silenced in melanoma cells by mechanisms triggered by TGF-beta and mediated by DNA methyltransferase 3 alpha (DNMT3A), which play a functional role in the epithelial-mesenchymal transition process and metastasis. In addition, TET2 (and TET3) may be guardians of regulatory T cell stability and immune homeostasis. TET2 belongs to the TET/JBP family of dioxygenases that require Fe2+ and alpha-ketoglutarate (also known as 2-oxoglutarate) for activity.
:
Pssm-ID: 380675 Cd Length: 434 Bit Score: 903.19 E-value: 0e+00
oxygenase domain of ten-eleven translocation (TET)2 methylcytosine dioxygenase and similar ...
3-447
0e+00
oxygenase domain of ten-eleven translocation (TET)2 methylcytosine dioxygenase and similar proteins; TET2 is involved in DNA demethylation through iteratively oxidizing 5-methylcytosine (5mC) into 5-hydroxymethylcytosine (5hmC), 5-formylcytosine (5fC) and 5-carboxylcytosine (5caC). Human TET2 (and TET1) have been shown to be more active on 5mC-DNA than 5hmC/5fC-DNA substrates. TET proteins contain a C-terminal catalytic domain which consists of a cysteine-rich region and a double-stranded beta-helix (DSBH) fold. TET2 acts as a tumor suppressor in hematopoiesis; mutations/deletions of the TET2 gene frequently occur in multiple spectra of myeloid malignancies. TET2 (and TET3) promoters are silenced in melanoma cells by mechanisms triggered by TGF-beta and mediated by DNA methyltransferase 3 alpha (DNMT3A), which play a functional role in the epithelial-mesenchymal transition process and metastasis. In addition, TET2 (and TET3) may be guardians of regulatory T cell stability and immune homeostasis. TET2 belongs to the TET/JBP family of dioxygenases that require Fe2+ and alpha-ketoglutarate (also known as 2-oxoglutarate) for activity.
Pssm-ID: 380675 Cd Length: 434 Bit Score: 903.19 E-value: 0e+00
Oxygenase domain of the 2OGFeDO superfamily; A double-stranded beta helix (DSBH) fold domain ...
164-431
7.43e-71
Oxygenase domain of the 2OGFeDO superfamily; A double-stranded beta helix (DSBH) fold domain of the 2-oxoglutarate (2OG)-Fe(II)-dependent dioxygenase (2OGFeDO) superfamily found in various eukaryotes, bacteria and bacteriophages. Members of this family catalyze nucleic acid modifications, such as thymidine hydroxylation during base J synthesis in kinetoplastids, and the conversion of 5 methyl-cytosine (5-mC) to 5-hydroxymethyl-cytosine (hmC), or further oxidation to 5-formylcytosine (5fC) and 5-carboxylcytosine (5caC). Metazoan TET proteins contain a cysteine-rich region inserted into the core of the DSBH fold. Vertebrate TET proteins are oncogenes that are mutated in various myeloid cancers. Fungal and algal versions of this family are linked to a predicted transposase and show lineage-specific expansions.
Pssm-ID: 372343 Cd Length: 166 Bit Score: 221.87 E-value: 7.43e-71
oxygenase domain of ten-eleven translocation (TET)2 methylcytosine dioxygenase and similar ...
3-447
0e+00
oxygenase domain of ten-eleven translocation (TET)2 methylcytosine dioxygenase and similar proteins; TET2 is involved in DNA demethylation through iteratively oxidizing 5-methylcytosine (5mC) into 5-hydroxymethylcytosine (5hmC), 5-formylcytosine (5fC) and 5-carboxylcytosine (5caC). Human TET2 (and TET1) have been shown to be more active on 5mC-DNA than 5hmC/5fC-DNA substrates. TET proteins contain a C-terminal catalytic domain which consists of a cysteine-rich region and a double-stranded beta-helix (DSBH) fold. TET2 acts as a tumor suppressor in hematopoiesis; mutations/deletions of the TET2 gene frequently occur in multiple spectra of myeloid malignancies. TET2 (and TET3) promoters are silenced in melanoma cells by mechanisms triggered by TGF-beta and mediated by DNA methyltransferase 3 alpha (DNMT3A), which play a functional role in the epithelial-mesenchymal transition process and metastasis. In addition, TET2 (and TET3) may be guardians of regulatory T cell stability and immune homeostasis. TET2 belongs to the TET/JBP family of dioxygenases that require Fe2+ and alpha-ketoglutarate (also known as 2-oxoglutarate) for activity.
Pssm-ID: 380675 Cd Length: 434 Bit Score: 903.19 E-value: 0e+00
oxygenase domain of ten-eleven translocation (TET)1, TET2, and TET3 methylcytosine ...
7-447
0e+00
oxygenase domain of ten-eleven translocation (TET)1, TET2, and TET3 methylcytosine dioxygenases and similar proteins; TET proteins are involved in DNA demethylation through iteratively oxidizing 5-methylcytosine (5mC) into 5-hydroxymethylcytosine (5hmC), 5-formylcytosine (5fC) and 5-carboxylcytosine (5caC). TET proteins contain a C-terminal catalytic domain which consists of a cysteine-rich region and a double-stranded beta-helix (DSBH) fold. Alterations in TET protein function have been linked to cancer, and TETs influence many cell differentiation processes. TET family genes have been implicated as tumor suppressors, for example mutations/deletions of the TET2 gene frequently occur in multiple spectra of myeloid malignancies. TET3 acts as a suppressor of ovarian cancer by demethylating the miR-30d precursor gene promoter to block TGF-beta1 induced epithelial-mesenchymal transition (EMT). TET3 (and TET2) promoters are silenced in melanoma cells by mechanisms triggered by TGF-beta and mediated by DNA methyltransferase 3 alpha (DNMT3A). TET genes are downregulated in endometriosis. TET proteins belong to the TET/JBP family of dioxygenases that require Fe2+ and alpha-ketoglutarate (also known as 2-oxoglutarate) for activity.
Pssm-ID: 380671 Cd Length: 398 Bit Score: 769.92 E-value: 0e+00
oxygenase domain of ten-eleven translocation (TET)1 methylcytosine dioxygenase and similar ...
7-447
0e+00
oxygenase domain of ten-eleven translocation (TET)1 methylcytosine dioxygenase and similar proteins; TET1 is involved in DNA demethylation through iteratively oxidizing 5-methylcytosine (5mC) into 5-hydroxymethylcytosine (5hmC), 5-formylcytosine (5fC) and 5-carboxylcytosine (5caC). Human TET1 (and TET2) are more active on 5mC-DNA than 5hmC/5fC-DNA substrates. TET proteins contain a C-terminal catalytic domain which consists of a cysteine-rich region and a double-stranded beta-helix (DSBH) fold. TET1 plays multiple roles in in tumor development and progression. TET1 serves as a tumor suppressor gene; loss of TET1 is associated with tumorigenesis and can be used as a potential biomarker for cancer therapy. In addition to its dioxygenase activity, it can induce epithelial-mesenchymal transition and act as a coactivator to regulate gene transcription. The regulation of TET1 is also correlated with microRNA in a posttranscriptional modification process. TET1 belongs to the TET/JBP family of dioxygenases that require Fe2+ and alpha-ketoglutarate (also known as 2-oxoglutarate) for activity.
Pssm-ID: 380674 Cd Length: 410 Bit Score: 754.44 E-value: 0e+00
oxygenase domain of ten-eleven translocation (TET)3 methylcytosine dioxygenase and similar ...
7-447
0e+00
oxygenase domain of ten-eleven translocation (TET)3 methylcytosine dioxygenase and similar proteins; TET3 is involved in DNA demethylation through iteratively oxidizing 5-methylcytosine (5mC) into 5-hydroxymethylcytosine (5hmC), 5-formylcytosine (5fC) and 5-carboxylcytosine (5caC). TET proteins contain a C-terminal catalytic domain which consists of a cysteine-rich region and a double-stranded beta-helix (DSBH) fold. TET3 serves as a tumor suppressor; it acts as a suppressor of ovarian cancer by demethylating the miR-30d precursor gene promoter to block TGF-beta1 induced epithelial-mesenchymal transition (EMT). TET3 (and TET2) promoters are silenced in melanoma cells by mechanisms triggered by TGF-beta and mediated by DNA methyltransferase 3 alpha (DNMT3A), which play a functional role in the EMT process and metastasis. In addition, TET3 (and TET2) may be guardians of regulatory T cell stability and immune homeostasis. TET3 has been shown to prevent terminal differentiation of adult neural stem cells by a mechanism involving direct binding and repression of TET3 to the imprinted gene Snrpn. TET3 has also been shown to mediate the activation of hepatic stellate cells via modulation of the long non-coding RNA HIF1A-AS1 expression. TET1 belongs to the TET/JBP family of dioxygenases that require Fe2+ and alpha-ketoglutarate (also known as 2-oxoglutarate) for activity.
Pssm-ID: 380676 Cd Length: 452 Bit Score: 716.77 E-value: 0e+00
oxygenase domain of ten-eleven translocation (TET) enzymes, J-binding proteins (JBPs), and ...
34-431
1.05e-101
oxygenase domain of ten-eleven translocation (TET) enzymes, J-binding proteins (JBPs), and similar proteins; TET proteins are involved in DNA demethylation through iteratively oxidizing 5-methylcytosine (5mC) into 5-hydroxymethylcytosine (5hmC), 5-formylcytosine (5fC) and 5-carboxylcytosine (5caC). TET proteins contain a C-terminal catalytic domain which consists of a cysteine-rich region and a double-stranded beta-helix (DSBH) fold. Alterations in TET protein function have been linked to cancer, and TETs influence many cell differentiation processes. J binding protein (JBP) 1 and JBP2 are thymidine hydroxylases that catalyze the first step of base J biosynthesis: the hydroxylation of thymine in DNA to form 5-hydroxymethyluracil (hmU). Base J (beta-d-glucopyranosyloxymethyluracil) is a hyper-modified DNA base found in the DNA of kinetoplastids (Trypanosoma brucei, Trypanosoma cruzi, and Leishmania). JBP1 and JBP2 each contain a J-DNA binding domain and a thymidine hydroxylase domain. Members of this TET/JBP family of dioxygenases require Fe2+ and alpha-ketoglutarate (also known as 2-oxoglutarate) for activity.
Pssm-ID: 380670 Cd Length: 264 Bit Score: 304.69 E-value: 1.05e-101
Oxygenase domain of the 2OGFeDO superfamily; A double-stranded beta helix (DSBH) fold domain ...
164-431
7.43e-71
Oxygenase domain of the 2OGFeDO superfamily; A double-stranded beta helix (DSBH) fold domain of the 2-oxoglutarate (2OG)-Fe(II)-dependent dioxygenase (2OGFeDO) superfamily found in various eukaryotes, bacteria and bacteriophages. Members of this family catalyze nucleic acid modifications, such as thymidine hydroxylation during base J synthesis in kinetoplastids, and the conversion of 5 methyl-cytosine (5-mC) to 5-hydroxymethyl-cytosine (hmC), or further oxidation to 5-formylcytosine (5fC) and 5-carboxylcytosine (5caC). Metazoan TET proteins contain a cysteine-rich region inserted into the core of the DSBH fold. Vertebrate TET proteins are oncogenes that are mutated in various myeloid cancers. Fungal and algal versions of this family are linked to a predicted transposase and show lineage-specific expansions.
Pssm-ID: 372343 Cd Length: 166 Bit Score: 221.87 E-value: 7.43e-71
Database: CDSEARCH/cdd Low complexity filter: no Composition Based Adjustment: yes E-value threshold: 0.01
References:
Wang J et al. (2023), "The conserved domain database in 2023", Nucleic Acids Res.51(D)384-8.
Lu S et al. (2020), "The conserved domain database in 2020", Nucleic Acids Res.48(D)265-8.
Marchler-Bauer A et al. (2017), "CDD/SPARCLE: functional classification of proteins via subfamily domain architectures.", Nucleic Acids Res.45(D)200-3.
of the residues that compose this conserved feature have been mapped to the query sequence.
Click on the triangle to view details about the feature, including a multiple sequence alignment
of your query sequence and the protein sequences used to curate the domain model,
where hash marks (#) above the aligned sequences show the location of the conserved feature residues.
The thumbnail image, if present, provides an approximate view of the feature's location in 3 dimensions.
Click on the triangle for interactive 3D structure viewing options.
Functional characterization of the conserved domain architecture found on the query.
Click here to see more details.
This image shows a graphical summary of conserved domains identified on the query sequence.
The Show Concise/Full Display button at the top of the page can be used to select the desired level of detail: only top scoring hits
(labeled illustration) or all hits
(labeled illustration).
Domains are color coded according to superfamilies
to which they have been assigned. Hits with scores that pass a domain-specific threshold
(specific hits) are drawn in bright colors.
Others (non-specific hits) and
superfamily placeholders are drawn in pastel colors.
if a domain or superfamily has been annotated with functional sites (conserved features),
they are mapped to the query sequence and indicated through sets of triangles
with the same color and shade of the domain or superfamily that provides the annotation. Mouse over the colored bars or triangles to see descriptions of the domains and features.
click on the bars or triangles to view your query sequence embedded in a multiple sequence alignment of the proteins used to develop the corresponding domain model.
The table lists conserved domains identified on the query sequence. Click on the plus sign (+) on the left to display full descriptions, alignments, and scores.
Click on the domain model's accession number to view the multiple sequence alignment of the proteins used to develop the corresponding domain model.
To view your query sequence embedded in that multiple sequence alignment, click on the colored bars in the Graphical Summary portion of the search results page,
or click on the triangles, if present, that represent functional sites (conserved features)
mapped to the query sequence.
Concise Display shows only the best scoring domain model, in each hit category listed below except non-specific hits, for each region on the query sequence.
(labeled illustration) Standard Display shows only the best scoring domain model from each source, in each hit category listed below for each region on the query sequence.
(labeled illustration) Full Display shows all domain models, in each hit category below, that meet or exceed the RPS-BLAST threshold for statistical significance.
(labeled illustration) Four types of hits can be shown, as available,
for each region on the query sequence:
specific hits meet or exceed a domain-specific e-value threshold
(illustrated example)
and represent a very high confidence that the query sequence belongs to the same protein family as the sequences use to create the domain model
non-specific hits
meet or exceed the RPS-BLAST threshold for statistical significance (default E-value cutoff of 0.01, or an E-value selected by user via the
advanced search options)
the domain superfamily to which the specific and non-specific hits belong
multi-domain models that were computationally detected and are likely to contain multiple single domains
Retrieve proteins that contain one or more of the domains present in the query sequence, using the Conserved Domain Architecture Retrieval Tool
(CDART).
Modify your query to search against a different database and/or use advanced search options