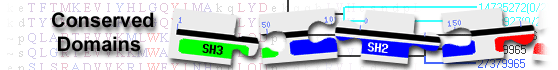


exonuclease 1, isoform CRA_c [Homo sapiens]
Protein Classification

exonuclease 1( domain architecture ID 10177090)
exonuclease 1 is a 5'->3' double-stranded DNA exonuclease that could act in a pathway that corrects mismatched base pairs; may also possess a cryptic 3'->5' double-stranded DNA exonuclease activity
List of domain hits
| Name | Accession | Description | Interval | E-value | ||||
| PIN_EXO1 | cd09857 | FEN-like PIN domains of Exonuclease-1, a structure-specific, divalent-metal-ion dependent, 5' ... |
1-201 | 1.25e-124 | ||||
FEN-like PIN domains of Exonuclease-1, a structure-specific, divalent-metal-ion dependent, 5' nuclease and homologs; exonuclease-1 (EXO1) is involved in multiple, eukaryotic DNA metabolic pathways, including DNA replication processes (5' flap DNA endonuclease activity and double stranded DNA 5'-exonuclease activity), DNA repair processes (DNA mismatch repair (MMR) and post-replication repair (PRR)), recombination, and telomere integrity. EXO1 functions in the MMS2 error-free branch of the PRR pathway in the maintenance and repair of stalled replication forks. Studies also suggest that EXO1 plays both structural and catalytic roles during MMR-mediated mutation avoidance. These nucleases are members of the structure-specific, 5' nuclease family (FEN-like) that catalyzes hydrolysis of DNA duplex-containing nucleic acid structures during DNA replication, repair, and recombination. Canonical members of the FEN-like family possess a PIN domain with a two-helical structure insert (also known as the helical arch, helical clamp or I domain) of variable length (approximately 16 to 800 residues), and at the C-terminus of the PIN domain a H3TH (helix-3-turn-helix) domain, an atypical helix-hairpin-helix-2-like region. Both the H3TH domain (not included in this model) and the helical arch/clamp region are involved in DNA binding. The PIN domain belongs to a large nuclease superfamily. The structural properties of the PIN domain indicate its putative active center, consisting of invariant acidic amino acid residues (putative metal-binding residues), is geometrically similar in the active center of structure-specific 5' nucleases, PIN-domain ribonucleases of eukaryotic rRNA editing proteins, and bacterial toxins of toxin-antitoxin (TA) operons. EXO1 nucleases also have C-terminal Mlh1- and Msh2-binding domains which allow interaction with MMR and PRR proteins, respectively. : Pssm-ID: 350207 [Multi-domain] Cd Length: 202 Bit Score: 371.74 E-value: 1.25e-124
|
||||||||
| H3TH_EXO1 | cd09908 | H3TH domain of Exonuclease-1, a structure-specific, divalent-metal-ion dependent, 5' nuclease; ... |
213-289 | 7.27e-35 | ||||
H3TH domain of Exonuclease-1, a structure-specific, divalent-metal-ion dependent, 5' nuclease; Exonuclease-1 (EXO1) is involved in multiple, eukaryotic DNA metabolic pathways, including DNA replication processes (5' flap DNA endonuclease activity and double stranded DNA 5'-exonuclease activity), DNA repair processes (DNA mismatch repair (MMR) and post-replication repair (PRR), recombination, and telomere integrity. EXO1 functions in the MMS2 error-free branch of the PRR pathway in the maintenance and repair of stalled replication forks. Studies also suggest that EXO1 plays both structural and catalytic roles during MMR-mediated mutation avoidance. Members of this subgroup include the H3TH (helix-3-turn-helix) domains of EXO1 and other similar eukaryotic 5' nucleases. These nucleases contain a PIN (PilT N terminus) domain with a helical arch/clamp region/I domain (not included here) and inserted within the PIN domain is an atypical helix-hairpin-helix-2 (HhH2)-like region. This atypical HhH2 region, the H3TH domain, has an extended loop with at least three turns between the first two helices, and only three of the four helices appear to be conserved. Both the H3TH domain and the helical arch/clamp region are involved in DNA binding. Studies suggest that a glycine-rich loop in the H3TH domain contacts the phosphate backbone of the template strand in the downstream DNA duplex. These nucleases have a carboxylate rich active site that is involved in binding essential divalent metal ion cofactors (Mg2+ or Mn2+) required for nuclease activity. The first metal binding site is composed entirely of Asp/Glu residues from the PIN domain, whereas, the second metal binding site is composed generally of two Asp residues from the PIN domain and one Asp residue from the H3TH domain. Together with the helical arch and network of amino acids interacting with metal binding ions, the H3TH region defines a positively charged active-site DNA-binding groove in structure-specific 5' nucleases. EXO1 nucleases also have C-terminal Mlh1- and Msh2-binding domains which allow interaction with MMR and PRR proteins, respectively. : Pssm-ID: 188628 [Multi-domain] Cd Length: 73 Bit Score: 126.92 E-value: 7.27e-35
|
||||||||
| Name | Accession | Description | Interval | E-value | ||||||
| PIN_EXO1 | cd09857 | FEN-like PIN domains of Exonuclease-1, a structure-specific, divalent-metal-ion dependent, 5' ... |
1-201 | 1.25e-124 | ||||||
FEN-like PIN domains of Exonuclease-1, a structure-specific, divalent-metal-ion dependent, 5' nuclease and homologs; exonuclease-1 (EXO1) is involved in multiple, eukaryotic DNA metabolic pathways, including DNA replication processes (5' flap DNA endonuclease activity and double stranded DNA 5'-exonuclease activity), DNA repair processes (DNA mismatch repair (MMR) and post-replication repair (PRR)), recombination, and telomere integrity. EXO1 functions in the MMS2 error-free branch of the PRR pathway in the maintenance and repair of stalled replication forks. Studies also suggest that EXO1 plays both structural and catalytic roles during MMR-mediated mutation avoidance. These nucleases are members of the structure-specific, 5' nuclease family (FEN-like) that catalyzes hydrolysis of DNA duplex-containing nucleic acid structures during DNA replication, repair, and recombination. Canonical members of the FEN-like family possess a PIN domain with a two-helical structure insert (also known as the helical arch, helical clamp or I domain) of variable length (approximately 16 to 800 residues), and at the C-terminus of the PIN domain a H3TH (helix-3-turn-helix) domain, an atypical helix-hairpin-helix-2-like region. Both the H3TH domain (not included in this model) and the helical arch/clamp region are involved in DNA binding. The PIN domain belongs to a large nuclease superfamily. The structural properties of the PIN domain indicate its putative active center, consisting of invariant acidic amino acid residues (putative metal-binding residues), is geometrically similar in the active center of structure-specific 5' nucleases, PIN-domain ribonucleases of eukaryotic rRNA editing proteins, and bacterial toxins of toxin-antitoxin (TA) operons. EXO1 nucleases also have C-terminal Mlh1- and Msh2-binding domains which allow interaction with MMR and PRR proteins, respectively. Pssm-ID: 350207 [Multi-domain] Cd Length: 202 Bit Score: 371.74 E-value: 1.25e-124
|
||||||||||
| H3TH_EXO1 | cd09908 | H3TH domain of Exonuclease-1, a structure-specific, divalent-metal-ion dependent, 5' nuclease; ... |
213-289 | 7.27e-35 | ||||||
H3TH domain of Exonuclease-1, a structure-specific, divalent-metal-ion dependent, 5' nuclease; Exonuclease-1 (EXO1) is involved in multiple, eukaryotic DNA metabolic pathways, including DNA replication processes (5' flap DNA endonuclease activity and double stranded DNA 5'-exonuclease activity), DNA repair processes (DNA mismatch repair (MMR) and post-replication repair (PRR), recombination, and telomere integrity. EXO1 functions in the MMS2 error-free branch of the PRR pathway in the maintenance and repair of stalled replication forks. Studies also suggest that EXO1 plays both structural and catalytic roles during MMR-mediated mutation avoidance. Members of this subgroup include the H3TH (helix-3-turn-helix) domains of EXO1 and other similar eukaryotic 5' nucleases. These nucleases contain a PIN (PilT N terminus) domain with a helical arch/clamp region/I domain (not included here) and inserted within the PIN domain is an atypical helix-hairpin-helix-2 (HhH2)-like region. This atypical HhH2 region, the H3TH domain, has an extended loop with at least three turns between the first two helices, and only three of the four helices appear to be conserved. Both the H3TH domain and the helical arch/clamp region are involved in DNA binding. Studies suggest that a glycine-rich loop in the H3TH domain contacts the phosphate backbone of the template strand in the downstream DNA duplex. These nucleases have a carboxylate rich active site that is involved in binding essential divalent metal ion cofactors (Mg2+ or Mn2+) required for nuclease activity. The first metal binding site is composed entirely of Asp/Glu residues from the PIN domain, whereas, the second metal binding site is composed generally of two Asp residues from the PIN domain and one Asp residue from the H3TH domain. Together with the helical arch and network of amino acids interacting with metal binding ions, the H3TH region defines a positively charged active-site DNA-binding groove in structure-specific 5' nucleases. EXO1 nucleases also have C-terminal Mlh1- and Msh2-binding domains which allow interaction with MMR and PRR proteins, respectively. Pssm-ID: 188628 [Multi-domain] Cd Length: 73 Bit Score: 126.92 E-value: 7.27e-35
|
||||||||||
| PTZ00217 | PTZ00217 | flap endonuclease-1; Provisional |
1-306 | 8.37e-33 | ||||||
flap endonuclease-1; Provisional Pssm-ID: 240317 [Multi-domain] Cd Length: 393 Bit Score: 131.28 E-value: 8.37e-33
|
||||||||||
| XPGN | smart00485 | Xeroderma pigmentosum G N-region; domain in nucleases |
1-86 | 2.12e-30 | ||||||
Xeroderma pigmentosum G N-region; domain in nucleases Pssm-ID: 214690 [Multi-domain] Cd Length: 99 Bit Score: 115.02 E-value: 2.12e-30
|
||||||||||
| XPG_I | pfam00867 | XPG I-region; |
141-225 | 4.96e-29 | ||||||
XPG I-region; Pssm-ID: 459970 [Multi-domain] Cd Length: 87 Bit Score: 110.68 E-value: 4.96e-29
|
||||||||||
| rad2 | TIGR00600 | DNA excision repair protein (rad2); All proteins in this family for which functions are known ... |
97-242 | 1.16e-06 | ||||||
DNA excision repair protein (rad2); All proteins in this family for which functions are known are flap endonucleases that generate the 3' incision next to DNA damage as part of nucleotide excision repair. This family is related to many other flap endonuclease families including the fen1 family. This family is based on the phylogenomic analysis of JA Eisen (1999, Ph.D. Thesis, Stanford University). [DNA metabolism, DNA replication, recombination, and repair] Pssm-ID: 273166 [Multi-domain] Cd Length: 1034 Bit Score: 52.21 E-value: 1.16e-06
|
||||||||||
| HhH2 | smart00279 | Helix-hairpin-helix class 2 (Pol1 family) motifs; |
213-243 | 1.59e-04 | ||||||
Helix-hairpin-helix class 2 (Pol1 family) motifs; Pssm-ID: 197623 [Multi-domain] Cd Length: 36 Bit Score: 39.35 E-value: 1.59e-04
|
||||||||||
| Name | Accession | Description | Interval | E-value | ||||||
| PIN_EXO1 | cd09857 | FEN-like PIN domains of Exonuclease-1, a structure-specific, divalent-metal-ion dependent, 5' ... |
1-201 | 1.25e-124 | ||||||
FEN-like PIN domains of Exonuclease-1, a structure-specific, divalent-metal-ion dependent, 5' nuclease and homologs; exonuclease-1 (EXO1) is involved in multiple, eukaryotic DNA metabolic pathways, including DNA replication processes (5' flap DNA endonuclease activity and double stranded DNA 5'-exonuclease activity), DNA repair processes (DNA mismatch repair (MMR) and post-replication repair (PRR)), recombination, and telomere integrity. EXO1 functions in the MMS2 error-free branch of the PRR pathway in the maintenance and repair of stalled replication forks. Studies also suggest that EXO1 plays both structural and catalytic roles during MMR-mediated mutation avoidance. These nucleases are members of the structure-specific, 5' nuclease family (FEN-like) that catalyzes hydrolysis of DNA duplex-containing nucleic acid structures during DNA replication, repair, and recombination. Canonical members of the FEN-like family possess a PIN domain with a two-helical structure insert (also known as the helical arch, helical clamp or I domain) of variable length (approximately 16 to 800 residues), and at the C-terminus of the PIN domain a H3TH (helix-3-turn-helix) domain, an atypical helix-hairpin-helix-2-like region. Both the H3TH domain (not included in this model) and the helical arch/clamp region are involved in DNA binding. The PIN domain belongs to a large nuclease superfamily. The structural properties of the PIN domain indicate its putative active center, consisting of invariant acidic amino acid residues (putative metal-binding residues), is geometrically similar in the active center of structure-specific 5' nucleases, PIN-domain ribonucleases of eukaryotic rRNA editing proteins, and bacterial toxins of toxin-antitoxin (TA) operons. EXO1 nucleases also have C-terminal Mlh1- and Msh2-binding domains which allow interaction with MMR and PRR proteins, respectively. Pssm-ID: 350207 [Multi-domain] Cd Length: 202 Bit Score: 371.74 E-value: 1.25e-124
|
||||||||||
| PIN_FEN1_EXO1-like | cd00128 | FEN-like PIN domains of Flap endonuclease-1 (FEN1)-like and exonuclease-1 (EXO1)-like ... |
6-202 | 8.97e-64 | ||||||
FEN-like PIN domains of Flap endonuclease-1 (FEN1)-like and exonuclease-1 (EXO1)-like nucleases, structure-specific, divalent-metal-ion dependent, 5' nucleases; PIN (PilT N terminus) domain of Flap endonuclease-1 (FEN1) and exonuclease-1 (EXO1)-like nucleases: FEN1, EXO1, Mkt1, Gap endonuclease 1 (GEN1) and Xeroderma pigmentosum complementation group G (XPG) nuclease. These nucleases are members of the structure-specific, 5' nuclease family (FEN-like) that catalyzes hydrolysis of DNA duplex-containing nucleic acid structures during DNA replication, repair, and recombination. Canonical members of the FEN-like family possess a PIN domain with a two-helical structure insert (also known as the helical arch, helical clamp or I domain) of variable length (approximately 16 to 800 residues), and at the C-terminus of the PIN domain a H3TH (helix-3-turn-helix) domain, an atypical helix-hairpin-helix-2-like region. Both the H3TH domain (not included in this model) and the helical arch/clamp region are involved in DNA binding. The PIN domain belongs to a large nuclease superfamily. The structural properties of the PIN domain indicate its putative active center, consisting of invariant acidic amino acid residues (putative metal-binding residues), is geometrically similar in the active center of structure-specific 5' nucleases, PIN-domain ribonucleases of eukaryotic rRNA editing proteins, and bacterial toxins of toxin-antitoxin (TA) operons. Pssm-ID: 350200 [Multi-domain] Cd Length: 162 Bit Score: 210.69 E-value: 8.97e-64
|
||||||||||
| H3TH_EXO1 | cd09908 | H3TH domain of Exonuclease-1, a structure-specific, divalent-metal-ion dependent, 5' nuclease; ... |
213-289 | 7.27e-35 | ||||||
H3TH domain of Exonuclease-1, a structure-specific, divalent-metal-ion dependent, 5' nuclease; Exonuclease-1 (EXO1) is involved in multiple, eukaryotic DNA metabolic pathways, including DNA replication processes (5' flap DNA endonuclease activity and double stranded DNA 5'-exonuclease activity), DNA repair processes (DNA mismatch repair (MMR) and post-replication repair (PRR), recombination, and telomere integrity. EXO1 functions in the MMS2 error-free branch of the PRR pathway in the maintenance and repair of stalled replication forks. Studies also suggest that EXO1 plays both structural and catalytic roles during MMR-mediated mutation avoidance. Members of this subgroup include the H3TH (helix-3-turn-helix) domains of EXO1 and other similar eukaryotic 5' nucleases. These nucleases contain a PIN (PilT N terminus) domain with a helical arch/clamp region/I domain (not included here) and inserted within the PIN domain is an atypical helix-hairpin-helix-2 (HhH2)-like region. This atypical HhH2 region, the H3TH domain, has an extended loop with at least three turns between the first two helices, and only three of the four helices appear to be conserved. Both the H3TH domain and the helical arch/clamp region are involved in DNA binding. Studies suggest that a glycine-rich loop in the H3TH domain contacts the phosphate backbone of the template strand in the downstream DNA duplex. These nucleases have a carboxylate rich active site that is involved in binding essential divalent metal ion cofactors (Mg2+ or Mn2+) required for nuclease activity. The first metal binding site is composed entirely of Asp/Glu residues from the PIN domain, whereas, the second metal binding site is composed generally of two Asp residues from the PIN domain and one Asp residue from the H3TH domain. Together with the helical arch and network of amino acids interacting with metal binding ions, the H3TH region defines a positively charged active-site DNA-binding groove in structure-specific 5' nucleases. EXO1 nucleases also have C-terminal Mlh1- and Msh2-binding domains which allow interaction with MMR and PRR proteins, respectively. Pssm-ID: 188628 [Multi-domain] Cd Length: 73 Bit Score: 126.92 E-value: 7.27e-35
|
||||||||||
| PTZ00217 | PTZ00217 | flap endonuclease-1; Provisional |
1-306 | 8.37e-33 | ||||||
flap endonuclease-1; Provisional Pssm-ID: 240317 [Multi-domain] Cd Length: 393 Bit Score: 131.28 E-value: 8.37e-33
|
||||||||||
| XPGN | smart00485 | Xeroderma pigmentosum G N-region; domain in nucleases |
1-86 | 2.12e-30 | ||||||
Xeroderma pigmentosum G N-region; domain in nucleases Pssm-ID: 214690 [Multi-domain] Cd Length: 99 Bit Score: 115.02 E-value: 2.12e-30
|
||||||||||
| XPG_I | pfam00867 | XPG I-region; |
141-225 | 4.96e-29 | ||||||
XPG I-region; Pssm-ID: 459970 [Multi-domain] Cd Length: 87 Bit Score: 110.68 E-value: 4.96e-29
|
||||||||||
| H3TH_FEN1-like | cd09901 | H3TH domains of Flap endonuclease-1 (FEN1)-like structure specific 5' nucleases: FEN1 ... |
213-289 | 8.48e-29 | ||||||
H3TH domains of Flap endonuclease-1 (FEN1)-like structure specific 5' nucleases: FEN1 (eukaryotic) and EXO1; The 5' nucleases within this family are capable of both 5'-3' exonucleolytic activity and cleaving bifurcated or branched DNA, in an endonucleolytic, structure-specific manner, and are involved in DNA replication, repair, and recombination. This family includes the H3TH (helix-3-turn-helix) domains of eukaryotic Flap Endonuclease-1 (FEN1), Exonuclease-1 (EXO1), and other eukaryotic homologs. These nucleases contain a PIN (PilT N terminus) domain with a helical arch/clamp region/I domain (not included here) and inserted within the PIN domain is an atypical helix-hairpin-helix-2 (HhH2)-like region. This atypical HhH2 region, the H3TH domain, has an extended loop with at least three turns between the first two helices, and only three of the four helices appear to be conserved. Both the H3TH domain and the helical arch/clamp region are involved in DNA binding. Studies suggest that a glycine-rich loop in the H3TH domain contacts the phosphate backbone of the template strand in the downstream DNA duplex. The nucleases within this family have a carboxylate rich active site that is involved in binding essential divalent metal ion cofactors (i. e., Mg2+, Mn2+, Zn2+, or Co2+) required for nuclease activity. The first metal binding site is composed entirely of Asp/Glu residues from the PIN domain, whereas, the second metal binding site is composed generally of two Asp residues from the PIN domain and one Asp residue from the H3TH domain. Together with the helical arch and network of amino acids interacting with metal binding ions, the H3TH region defines a positively charged active-site DNA-binding groove in structure-specific 5' nucleases. Pssm-ID: 188621 [Multi-domain] Cd Length: 73 Bit Score: 109.55 E-value: 8.48e-29
|
||||||||||
| PIN_XPG_RAD2 | cd09868 | FEN-like PIN domains of Xeroderma pigmentosum complementation group G (XPG) nuclease, a ... |
2-183 | 1.13e-25 | ||||||
FEN-like PIN domains of Xeroderma pigmentosum complementation group G (XPG) nuclease, a structure-specific, divalent-metal-ion dependent, 5' nuclease and homologs; The Xeroderma pigmentosum complementation group G (XPG) nuclease plays a central role in nucleotide excision repair (NER) in cleaving DNA bubble structures or loops. XPG is a member of the structure-specific, 5' nuclease family (FEN-like) that catalyzes hydrolysis of DNA duplex-containing nucleic acid structures during DNA replication, repair, and recombination. Canonical members of the FEN-like family possess a PIN domain with a two-helical structure insert (also known as the helical arch, helical clamp or I domain) of variable length (approximately 16 to 800 residues), and at the C-terminus of the PIN domain a H3TH (helix-3-turn-helix) domain, an atypical helix-hairpin-helix-2-like region. Both the H3TH domain (not included in this model) and the helical arch/clamp region are involved in DNA binding. The PIN domain belongs to a large nuclease superfamily. The structural properties of the PIN domain indicate its putative active center, consisting of invariant acidic amino acid residues (putative metal-binding residues), is geometrically similar in the active center of structure-specific 5' nucleases, PIN-domain ribonucleases of eukaryotic rRNA editing proteins, and bacterial toxins of toxin-antitoxin (TA) operons. Pssm-ID: 350216 [Multi-domain] Cd Length: 209 Bit Score: 105.29 E-value: 1.13e-25
|
||||||||||
| XPGI | smart00484 | Xeroderma pigmentosum G I-region; domain in nucleases |
138-207 | 2.61e-24 | ||||||
Xeroderma pigmentosum G I-region; domain in nucleases Pssm-ID: 214689 [Multi-domain] Cd Length: 73 Bit Score: 96.88 E-value: 2.61e-24
|
||||||||||
| PIN_FEN1-like | cd09856 | FEN-like PIN domains of Flap endonuclease-1 (FEN1)-like, structure-specific, ... |
5-183 | 1.64e-22 | ||||||
FEN-like PIN domains of Flap endonuclease-1 (FEN1)-like, structure-specific, divalent-metal-ion dependent, 5' nucleases; PIN (PilT N terminus) domain of Flap endonuclease-1 (FEN1)-like nucleases: FEN1, Gap endonuclease 1 (GEN1) and Xeroderma pigmentosum complementation group G (XPG) nuclease. Nucleases in this subfamily are members of the structure-specific, 5' nuclease family (FEN-like) that catalyzes hydrolysis of DNA duplex-containing nucleic acid structures during DNA replication, repair, and recombination. Canonical members of the FEN-like family possess a PIN domain with a two-helical structure insert (also known as the helical arch, helical clamp or I domain) of variable length (approximately 16 to 800 residues), and at the C-terminus of the PIN domain a H3TH (helix-3-turn-helix) domain, an atypical helix-hairpin-helix-2-like region. Both the H3TH domain (not included in this model) and the helical arch/clamp region are involved in DNA binding. The PIN domain belongs to a large nuclease superfamily. The structural properties of the PIN domain indicate its putative active center, consisting of invariant acidic amino acid residues (putative metal-binding residues), is geometrically similar in the active center of structure-specific 5' nucleases, PIN-domain ribonucleases of eukaryotic rRNA editing proteins, and bacterial toxins of toxin-antitoxin (TA) operons. Pssm-ID: 350206 [Multi-domain] Cd Length: 235 Bit Score: 97.22 E-value: 1.64e-22
|
||||||||||
| PIN_YEN1 | cd09870 | FEN-like PIN domains of Saccharomyces cerevisiae endonuclease 1 (YEN1), Chaetomium ... |
2-183 | 2.01e-22 | ||||||
FEN-like PIN domains of Saccharomyces cerevisiae endonuclease 1 (YEN1), Chaetomium thermophilum junction-resolving enzyme GEN1, and fungal homologs; Fungal Endonuclease 1 (YEN1 and GEN1, GEN1 is known as YEN1 in Saccharomyces cerevisiae) is a four-way (Holliday) junction resolvase. Members of this subgroup belong to the structure-specific, 5' nuclease family (FEN-like) that catalyzes hydrolysis of DNA duplex-containing nucleic acid structures during DNA replication, repair, and recombination. Canonical members of the FEN-like family possess a PIN domain with a two-helical structure insert (also known as the helical arch, helical clamp or I domain) of variable length (approximately 16 to 800 residues), and at the C-terminus of the PIN domain a H3TH (helix-3-turn-helix) domain, an atypical helix-hairpin-helix-2-like region. Both the H3TH domain (not included in this model) and the helical arch/clamp region are involved in DNA binding. The PIN domain belongs to a large nuclease superfamily. The structural properties of the PIN domain indicate its putative active center, consisting of invariant acidic amino acid residues (putative metal-binding residues), is geometrically similar in the active center of structure-specific 5' nucleases, PIN-domain ribonucleases of eukaryotic rRNA editing proteins, and bacterial toxins of toxin-antitoxin (TA) operons. Pssm-ID: 350218 [Multi-domain] Cd Length: 229 Bit Score: 96.57 E-value: 2.01e-22
|
||||||||||
| PIN_MKT1 | cd09858 | FEN-like PIN domains of Mkt1, a global regulator of mRNAs encoding mitochondrial proteins and ... |
61-187 | 8.62e-18 | ||||||
FEN-like PIN domains of Mkt1, a global regulator of mRNAs encoding mitochondrial proteins and eukaryotic homologs; The Mkt1 gene product interacts with the Poly(A)-binding protein associated factor, Pbp1, and is present at the 3' end of RNA transcripts during translation. The Mkt1-Pbp1 complex is involved in the post-transcriptional regulation of HO endonuclease expression. Mkt1 and eukaryotic homologs are atypical members of the structure-specific, 5' nuclease family (FEN-like). Canonical members of the FEN-like family possess a PIN domain with a two-helical structure insert (also known as the helical arch, helical clamp or I domain) of variable length (approximately 16 to 800 residues), and at the C-terminus of the PIN domain a H3TH (helix-3-turn-helix) domain, an atypical helix-hairpin-helix-2-like region. Both the H3TH domain (not included in this model) and the helical arch/clamp region are involved in DNA binding. Although Mkt1 appears to possess both a PIN and H3TH domain, the Mkt1 PIN domain lacks several of the active site residues necessary to bind essential divalent metal ion cofactors (Mg2+/Mn2+) required for nuclease activity in this family. Also, Mkt1 lacks the glycine-rich loop in the H3TH domain which is proposed to facilitate duplex DNA binding. Pssm-ID: 350208 [Multi-domain] Cd Length: 206 Bit Score: 82.58 E-value: 8.62e-18
|
||||||||||
| H3TH_FEN1-XPG-like | cd09897 | H3TH domains of Flap endonuclease-1 (FEN1)-like structure specific 5' nucleases; The 5' ... |
213-284 | 7.64e-17 | ||||||
H3TH domains of Flap endonuclease-1 (FEN1)-like structure specific 5' nucleases; The 5' nucleases within this family are capable of both 5'-3' exonucleolytic activity and cleaving bifurcated or branched DNA, in an endonucleolytic, structure-specific manner, and are involved in DNA replication, repair, and recombination. This family includes the H3TH (helix-3-turn-helix) domains of Flap Endonuclease-1 (FEN1), Exonuclease-1 (EXO1), Mkt1, Gap Endonuclease 1 (GEN1), Xeroderma pigmentosum complementation group G (XPG) nuclease, and other eukaryotic and archaeal homologs. These nucleases contain a PIN (PilT N terminus) domain with a helical arch/clamp region/I domain (not included here) and inserted within the PIN domain is an atypical helix-hairpin-helix-2 (HhH2)-like region. This atypical HhH2 region, the H3TH domain, has an extended loop with at least three turns between the first two helices, and only three of the four helices appear to be conserved. Both the H3TH domain and the helical arch/clamp region are involved in DNA binding. Studies suggest that a glycine-rich loop in the H3TH domain contacts the phosphate backbone of the template strand in the downstream DNA duplex. With the except of the Mkt1-like proteins, the nucleases within this family have a carboxylate rich active site that is involved in binding essential divalent metal ion cofactors (i. e., Mg2+, Mn2+, Zn2+, or Co2+) required for nuclease activity. The first metal binding site is composed entirely of Asp/Glu residues from the PIN domain, whereas, the second metal binding site is composed generally of two Asp residues from the PIN domain and one Asp residue from the H3TH domain. Together with the helical arch and network of amino acids interacting with metal binding ions, the H3TH region defines a positively charged active-site DNA-binding groove in structure-specific 5' nucleases. Pssm-ID: 188617 [Multi-domain] Cd Length: 68 Bit Score: 75.33 E-value: 7.64e-17
|
||||||||||
| XPG_N | pfam00752 | XPG N-terminal domain; |
2-86 | 2.27e-16 | ||||||
XPG N-terminal domain; Pssm-ID: 395609 [Multi-domain] Cd Length: 100 Bit Score: 75.09 E-value: 2.27e-16
|
||||||||||
| PRK03980 | PRK03980 | flap endonuclease-1; Provisional |
47-308 | 1.01e-15 | ||||||
flap endonuclease-1; Provisional Pssm-ID: 235185 [Multi-domain] Cd Length: 292 Bit Score: 78.32 E-value: 1.01e-15
|
||||||||||
| PIN_FEN1 | cd09867 | FEN-like PIN domains of Flap endonuclease-1, a structure-specific, divalent-metal-ion ... |
22-183 | 6.25e-15 | ||||||
FEN-like PIN domains of Flap endonuclease-1, a structure-specific, divalent-metal-ion dependent, 5' nuclease and homologs; Flap endonuclease-1 (FEN1) is involved in multiple DNA metabolic pathways, including DNA replication processes (5' flap DNA endonuclease activity and double stranded DNA 5'-exonuclease activity) and DNA repair processes (long-patch base excision repair) in eukaryotes and archaea. Interaction between FEN1 and PCNA (Proliferating cell nuclear antigen) is an essential prerequisite to FEN1's DNA replication functionality and stimulates FEN1 nuclease activity by 10-50 fold. FEN1 belongs to the FEN1-EXO1-like subfamily of structure-specific, 5' nucleases (FEN-like family). Canonical members of the FEN-like family possess a PIN domain with a two-helical structure insert (also known as the helical arch, helical clamp or I domain) of variable length (approximately 16 to 800 residues), and at the C-terminus of the PIN domain a H3TH (helix-3-turn-helix) domain, an atypical helix-hairpin-helix-2-like region. Both the H3TH domain (not included in this model) and the helical arch/clamp region are involved in DNA binding. The PIN domain belongs to a large nuclease superfamily. The structural properties of the PIN domain indicate its putative active center, consisting of invariant acidic amino acid residues (putative metal-binding residues), is geometrically similar in the active center of structure-specific 5' nucleases, PIN-domain ribonucleases of eukaryotic rRNA editing proteins, and bacterial toxins of toxin-antitoxin (TA) operons. FEN1 has a C-terminal extension containing residues forming the consensus PIP-box - Qxx(M/L/I)xxF(Y/F) which serves to anchor FEN1 to PCNA. Pssm-ID: 350215 [Multi-domain] Cd Length: 251 Bit Score: 75.13 E-value: 6.25e-15
|
||||||||||
| PIN_GEN1 | cd09869 | FEN-like PIN domains of Gap Endonuclease 1, a structure-specific, divalent-metal-ion dependent, ... |
2-188 | 3.00e-14 | ||||||
FEN-like PIN domains of Gap Endonuclease 1, a structure-specific, divalent-metal-ion dependent, 5' nuclease and homologs; Gap Endonuclease 1 (GEN1) is a Holliday junction resolvase reported to symmetrically cleave Holliday junctions and allow religation without additional processing. GEN1 is a member of the structure-specific, 5' nuclease family (FEN-like) that catalyzes hydrolysis of DNA duplex-containing nucleic acid structures during DNA replication, repair, and recombination. Canonical members of the FEN-like family possess a PIN domain with a two-helical structure insert (also known as the helical arch, helical clamp or I domain) of variable length (approximately 16 to 800 residues), and at the C-terminus of the PIN domain a H3TH (helix-3-turn-helix) domain, an atypical helix-hairpin-helix-2-like region. Both the H3TH domain (not included in this model) and the helical arch/clamp region are involved in DNA binding. The PIN domain belongs to a large nuclease superfamily. The structural properties of the PIN domain indicate its putative active center, consisting of invariant acidic amino acid residues (putative metal-binding residues), is geometrically similar in the active center of structure-specific 5' nucleases, PIN-domain ribonucleases of eukaryotic rRNA editing proteins, and bacterial toxins of toxin-antitoxin (TA) operons. Pssm-ID: 350217 [Multi-domain] Cd Length: 227 Bit Score: 72.64 E-value: 3.00e-14
|
||||||||||
| H3TH_FEN1-Euk | cd09907 | H3TH domain of Flap Endonuclease-1, a structure-specific, divalent-metal-ion dependent, 5' ... |
213-289 | 4.07e-09 | ||||||
H3TH domain of Flap Endonuclease-1, a structure-specific, divalent-metal-ion dependent, 5' nuclease: Eukaryotic homologs; Members of this subgroup include the H3TH (helix-3-turn-helix) domains of eukaryotic Flap endonuclease-1 (FEN1), 5' nucleases. FEN1 is involved in multiple DNA metabolic pathways, including DNA replication processes (5' flap DNA endonuclease activity and double stranded DNA 5'-exonuclease activity) and DNA repair processes (long-patch base excision repair) in eukaryotes and archaea. Interaction between FEN1 and PCNA (Proliferating cell nuclear antigen) is an essential prerequisite to FEN1's DNA replication functionality and stimulates FEN1 nuclease activity by 10-50 fold. These nucleases contain a PIN (PilT N terminus) domain with a helical arch/clamp region/I domain (not included here) and inserted within the PIN domain is an atypical helix-hairpin-helix-2 (HhH2)-like region. This atypical HhH2 region, the H3TH domain, has an extended loop with at least three turns between the first two helices, and only three of the four helices appear to be conserved. Both the H3TH domain and the helical arch/clamp region are involved in DNA binding. Studies suggest that a glycine-rich loop in the H3TH domain contacts the phosphate backbone of the template strand in the downstream DNA duplex. The nucleases within this subfamily have a carboxylate rich active site that is involved in binding essential divalent metal ion cofactors (Mg2+ or Mn2+) required for nuclease activity. The first metal binding site is composed entirely of Asp/Glu residues from the PIN domain, whereas, the second metal binding site is composed generally of two Asp residues from the PIN domain and one Asp residue from the H3TH domain. Together with the helical arch and network of amino acids interacting with metal binding ions, the H3TH region defines a positively charged active-site DNA-binding groove in structure-specific 5' nucleases. Also, FEN1 has a C-terminal extension containing residues forming the consensus PIP-box - Qxx(M/L/I)xxF(Y/F) which serves to anchor FEN1 to PCNA. Pssm-ID: 188627 [Multi-domain] Cd Length: 70 Bit Score: 53.70 E-value: 4.07e-09
|
||||||||||
| PIN_FEN-like | cd09853 | FEN-like PIN domains of structure-specific 5' nucleases (or Flap endonuclease-1-like) involved ... |
66-186 | 1.83e-07 | ||||||
FEN-like PIN domains of structure-specific 5' nucleases (or Flap endonuclease-1-like) involved in DNA replication, repair, and recombination; Structure-specific 5' nucleases are capable of both 5'-3' exonucleolytic activity and cleaving bifurcated or branched DNA, in an endonucleolytic, structure-specific manner. The family includes the PIN (PilT N terminus) domains of Flap endonuclease-1 (FEN1), exonuclease-1 (EXO1), Mkt1, Gap Endonuclease 1 (GEN1), and Xeroderma pigmentosum complementation group G (XPG) nuclease. Also included are the PIN domains of the 5'-3' exonucleases of DNA polymerase I and single domain protein homologs, as well as, the bacteriophage T4- and T5-5' nucleases, and other homologs. Canonical members of this FEN-like family possess a PIN domain with a two-helical structure insert (also known as the helical arch, helical clamp or I domain) of variable length (approximately 16 to 800 residues), and at the C-terminus of the PIN domain a H3TH (helix-3-turn-helix) domain, an atypical helix-hairpin-helix-2-like region. Both the H3TH domain (not included in this model) and the helical arch/clamp region are involved in DNA binding. The PIN domain belongs to a large nuclease superfamily. The structural properties of the PIN domain indicate its putative active center, consisting of invariant acidic amino acid residues (putative metal-binding residues), is geometrically similar in the active center of structure-specific 5' nucleases, PIN-domain ribonucleases of eukaryotic rRNA editing proteins, and bacterial toxins of toxin-antitoxin (TA) operons. Pssm-ID: 350204 [Multi-domain] Cd Length: 174 Bit Score: 51.72 E-value: 1.83e-07
|
||||||||||
| rad2 | TIGR00600 | DNA excision repair protein (rad2); All proteins in this family for which functions are known ... |
97-242 | 1.16e-06 | ||||||
DNA excision repair protein (rad2); All proteins in this family for which functions are known are flap endonucleases that generate the 3' incision next to DNA damage as part of nucleotide excision repair. This family is related to many other flap endonuclease families including the fen1 family. This family is based on the phylogenomic analysis of JA Eisen (1999, Ph.D. Thesis, Stanford University). [DNA metabolism, DNA replication, recombination, and repair] Pssm-ID: 273166 [Multi-domain] Cd Length: 1034 Bit Score: 52.21 E-value: 1.16e-06
|
||||||||||
| H3TH_StructSpec-5'-nucleases | cd00080 | H3TH domains of structure-specific 5' nucleases (or flap endonuclease-1-like) involved in DNA ... |
213-278 | 1.54e-06 | ||||||
H3TH domains of structure-specific 5' nucleases (or flap endonuclease-1-like) involved in DNA replication, repair, and recombination; The 5' nucleases of this superfamily are capable of both 5'-3' exonucleolytic activity and cleaving bifurcated or branched DNA, in an endonucleolytic, structure-specific manner, and are involved in DNA replication, repair, and recombination. The superfamily includes the H3TH (helix-3-turn-helix) domains of Flap Endonuclease-1 (FEN1), Exonuclease-1 (EXO1), Mkt1, Gap Endonuclease 1 (GEN1) and Xeroderma pigmentosum complementation group G (XPG) nuclease. Also included are the H3TH domains of the 5'-3' exonucleases of DNA polymerase I and single domain protein homologs, as well as, the bacteriophage T4 RNase H, T5-5'nuclease, and other homologs. These nucleases contain a PIN (PilT N terminus) domain with a helical arch/clamp region/I domain (not included here) and inserted within the C-terminal region of the PIN domain is an atypical helix-hairpin-helix-2 (HhH2)-like region. This atypical HhH2 region, the H3TH domain, has an extended loop with at least three turns between the first two helices, and only three of the four helices appear to be conserved. Both the H3TH domain and the helical arch/clamp region are involved in DNA binding. Studies suggest that a glycine-rich loop in the H3TH domain contacts the phosphate backbone of the template strand in the downstream DNA duplex. Typically, the nucleases within this superfamily have a carboxylate rich active site that is involved in binding essential divalent metal ion cofactors (i. e., Mg2+, Mn2+, Zn2+, or Co2+) required for nuclease activity. The first metal binding site is composed entirely of Asp/Glu residues from the PIN domain, whereas, the second metal binding site is composed generally of two Asp residues from the PIN domain and one or two Asp residues from the H3TH domain. Together with the helical arch and network of amino acids interacting with metal binding ions, the H3TH region defines a positively charged active-site DNA-binding groove in structure-specific 5' nucleases. Pssm-ID: 188616 [Multi-domain] Cd Length: 71 Bit Score: 46.21 E-value: 1.54e-06
|
||||||||||
| H3TH_XPG-like | cd09900 | H3TH domains of Flap endonuclease-1 (FEN1)-like structure specific 5' nucleases: FEN1 ... |
213-262 | 1.40e-04 | ||||||
H3TH domains of Flap endonuclease-1 (FEN1)-like structure specific 5' nucleases: FEN1 (archaeal), GEN1, YEN1, and XPG; The 5' nucleases within this family are capable of both 5'-3' exonucleolytic activity and cleaving bifurcated or branched DNA, in an endonucleolytic, structure-specific manner, and are involved in DNA replication, repair, and recombination. This family includes the H3TH (helix-3-turn-helix) domains of archaeal Flap Endonuclease-1 (FEN1), Gap Endonuclease 1 (GEN1), Yeast Endonuclease 1 (YEN1), Xeroderma pigmentosum complementation group G (XPG) nuclease, and other eukaryotic and archaeal homologs. These nucleases contain a PIN (PilT N terminus) domain with a helical arch/clamp region/I domain (not included here) and inserted within the PIN domain is an atypical helix-hairpin-helix-2 (HhH2)-like region. This atypical HhH2 region, the H3TH domain, has an extended loop with at least three turns between the first two helices, and only three of the four helices appear to be conserved. Both the H3TH domain and the helical arch/clamp region are involved in DNA binding. Studies suggest that a glycine-rich loop in the H3TH domain contacts the phosphate backbone of the template strand in the downstream DNA duplex. With the except of the Mkt1-like proteins, the nucleases within this family have a carboxylate rich active site that is involved in binding essential divalent metal ion cofactors (i. e., Mg2+, Mn2+, Zn2+, or Co2+) required for nuclease activity. The first metal binding site is composed entirely of Asp/Glu residues from the PIN domain, whereas, the second metal binding site is composed generally of two Asp residues from the PIN domain and one Asp residue from the H3TH domain. Together with the helical arch and network of amino acids interacting with metal binding ions, the H3TH region defines a positively charged active-site DNA-binding groove in structure-specific 5' nucleases. Pssm-ID: 188620 [Multi-domain] Cd Length: 52 Bit Score: 40.16 E-value: 1.40e-04
|
||||||||||
| HhH2 | smart00279 | Helix-hairpin-helix class 2 (Pol1 family) motifs; |
213-243 | 1.59e-04 | ||||||
Helix-hairpin-helix class 2 (Pol1 family) motifs; Pssm-ID: 197623 [Multi-domain] Cd Length: 36 Bit Score: 39.35 E-value: 1.59e-04
|
||||||||||
| rad2 | TIGR00600 | DNA excision repair protein (rad2); All proteins in this family for which functions are known ... |
1-79 | 5.19e-03 | ||||||
DNA excision repair protein (rad2); All proteins in this family for which functions are known are flap endonucleases that generate the 3' incision next to DNA damage as part of nucleotide excision repair. This family is related to many other flap endonuclease families including the fen1 family. This family is based on the phylogenomic analysis of JA Eisen (1999, Ph.D. Thesis, Stanford University). [DNA metabolism, DNA replication, recombination, and repair] Pssm-ID: 273166 [Multi-domain] Cd Length: 1034 Bit Score: 40.65 E-value: 5.19e-03
|
||||||||||
| Blast search parameters | ||||
|
