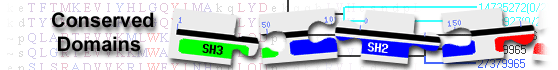


polyketide synthase dehydratase domain-containing protein, partial [Streptomyces vinaceusdrappus]
Protein Classification

List of domain hits
| Name | Accession | Description | Interval | E-value | |||||
| PS-DH | pfam14765 | Polyketide synthase dehydratase; This is the dehydratase domain of polyketide synthases. ... |
85-377 | 4.59e-52 | |||||
Polyketide synthase dehydratase; This is the dehydratase domain of polyketide synthases. Structural analysis shows these DH domains are double hotdogs in which the active site contains a histidine from the N-terminal hotdog and an aspartate from the C-terminal hotdog. Studies have uncovered that a substrate tunnel formed between the DH domains may be essential for loading substrates and unloading products. : Pssm-ID: 434191 Cd Length: 296 Bit Score: 179.49 E-value: 4.59e-52
|
|||||||||
| NADB_Rossmann super family | cl21454 | Rossmann-fold NAD(P)(+)-binding proteins; A large family of proteins that share a ... |
384-543 | 2.43e-28 | |||||
Rossmann-fold NAD(P)(+)-binding proteins; A large family of proteins that share a Rossmann-fold NAD(P)H/NAD(P)(+) binding (NADB) domain. The NADB domain is found in numerous dehydrogenases of metabolic pathways such as glycolysis, and many other redox enzymes. NAD binding involves numerous hydrogen-bonds and van der Waals contacts, in particular H-bonding of residues in a turn between the first strand and the subsequent helix of the Rossmann-fold topology. Characteristically, this turn exhibits a consensus binding pattern similar to GXGXXG, in which the first 2 glycines participate in NAD(P)-binding, and the third facilitates close packing of the helix to the beta-strand. Typically, proteins in this family contain a second domain in addition to the NADB domain, which is responsible for specifically binding a substrate and catalyzing a particular enzymatic reaction. The actual alignment was detected with superfamily member cd08956: Pssm-ID: 473865 [Multi-domain] Cd Length: 448 Bit Score: 117.75 E-value: 2.43e-28
|
|||||||||
| Name | Accession | Description | Interval | E-value | ||||||||
| PS-DH | pfam14765 | Polyketide synthase dehydratase; This is the dehydratase domain of polyketide synthases. ... |
85-377 | 4.59e-52 | ||||||||
Polyketide synthase dehydratase; This is the dehydratase domain of polyketide synthases. Structural analysis shows these DH domains are double hotdogs in which the active site contains a histidine from the N-terminal hotdog and an aspartate from the C-terminal hotdog. Studies have uncovered that a substrate tunnel formed between the DH domains may be essential for loading substrates and unloading products. Pssm-ID: 434191 Cd Length: 296 Bit Score: 179.49 E-value: 4.59e-52
|
||||||||||||
| PKS_DH | smart00826 | Dehydratase domain in polyketide synthase (PKS) enzymes; |
85-259 | 2.56e-49 | ||||||||
Dehydratase domain in polyketide synthase (PKS) enzymes; Pssm-ID: 214837 Cd Length: 167 Bit Score: 167.79 E-value: 2.56e-49
|
||||||||||||
| PksD | COG3321 | Acyl transferase domain in polyketide synthase (PKS) enzymes [Secondary metabolites ... |
2-543 | 1.41e-29 | ||||||||
Acyl transferase domain in polyketide synthase (PKS) enzymes [Secondary metabolites biosynthesis, transport and catabolism]; Pssm-ID: 442550 [Multi-domain] Cd Length: 1386 Bit Score: 124.21 E-value: 1.41e-29
|
||||||||||||
| KR_3_FAS_SDR_x | cd08956 | beta-ketoacyl reductase (KR) domain of fatty acid synthase (FAS), subgroup 3, complex (x); ... |
384-543 | 2.43e-28 | ||||||||
beta-ketoacyl reductase (KR) domain of fatty acid synthase (FAS), subgroup 3, complex (x); Ketoreductase, a module of the multidomain polyketide synthase (PKS), has 2 subdomains, each corresponding to a SDR family monomer. The C-terminal subdomain catalyzes the NADPH-dependent reduction of the beta-carbonyl of a polyketide to a hydroxyl group, a step in the biosynthesis of polyketides, such as erythromycin. The N-terminal subdomain, an interdomain linker, is a truncated Rossmann fold which acts to stabilizes the catalytic subdomain. Unlike typical SDRs, the isolated domain does not oligomerize but is composed of 2 subdomains, each resembling an SDR monomer. The active site resembles that of typical SDRs, except that the usual positions of the catalytic Asn and Tyr are swapped, so that the canonical YXXXK motif changes to YXXXN. Modular PKSs are multifunctional structures in which the makeup recapitulates that found in (and may have evolved from) FAS. In some instances, such as porcine FAS, an enoyl reductase (ER) module is inserted between the sub-domains. Fatty acid synthesis occurs via the stepwise elongation of a chain (which is attached to acyl carrier protein, ACP) with 2-carbon units. Eukaryotic systems consists of large, multifunctional synthases (type I) while bacterial, type II systems, use single function proteins. Fungal fatty acid synthesis uses a dodecamer of 6 alpha and 6 beta subunits. In mammalian type FAS cycles, ketoacyl synthase forms acetoacetyl-ACP which is reduced by the NADP-dependent beta-KR, forming beta-hydroxyacyl-ACP, which is in turn dehydrated by dehydratase to a beta-enoyl intermediate, which is reduced by NADP-dependent beta- ER. Polyketide synthesis also proceeds via the addition of 2-carbon units as in fatty acid synthesis. The complex SDR NADP-binding motif, GGXGXXG, is often present, but is not strictly conserved in each instance of the module. This subfamily includes KR domains found in many multidomain PKSs, including six of seven Sorangium cellulosum PKSs (encoded by spiDEFGHIJ) which participate in the synthesis of the polyketide scaffold of the cytotoxic spiroketal polyketide spirangien. These seven PKSs have either a single PKS module (SpiF), two PKR modules (SpiD,-E,-I,-J), or three PKS modules (SpiG,-H). This subfamily includes the second KR domains of SpiE,-G, I, and -J, both KR domains of SpiD, and the third KR domain of SpiH. The single KR domain of SpiF, the first and second KR domains of SpiH, the first KR domains of SpiE,-G,- I, and -J, and the third KR domain of SpiG, belong to a different KR_FAS_SDR subfamily. SDRs are a functionally diverse family of oxidoreductases that have a single domain with a structurally conserved Rossmann fold (alpha/beta folding pattern with a central beta-sheet), an NAD(P)(H)-binding region, and a structurally diverse C-terminal region. Classical SDRs are typically about 250 residues long, while extended SDRs are approximately 350 residues. Sequence identity between different SDR enzymes are typically in the 15-30% range, but the enzymes share the Rossmann fold NAD-binding motif and characteristic NAD-binding and catalytic sequence patterns. These enzymes catalyze a wide range of activities including the metabolism of steroids, cofactors, carbohydrates, lipids, aromatic compounds, and amino acids, and act in redox sensing. Classical SDRs have an TGXXX[AG]XG cofactor binding motif and a YXXXK active site motif, with the Tyr residue of the active site motif serving as a critical catalytic residue (Tyr-151, human prostaglandin dehydrogenase (PGDH) numbering). In addition to the Tyr and Lys, there is often an upstream Ser (Ser-138, PGDH numbering) and/or an Asn (Asn-107, PGDH numbering) contributing to the active site; while substrate binding is in the C-terminal region, which determines specificity. The standard reaction mechanism is a 4-pro-S hydride transfer and proton relay involving the conserved Tyr and Lys, a water molecule stabilized by Asn, and nicotinamide. Extended SDRs have additional elements in the C-terminal region, and typically have a TGXXGXXG cofactor binding motif. Complex (multidomain) SDRs such as ketoreductase domains of fatty acid synthase have a GGXGXXG NAD(P)-binding motif and an altered active site motif (YXXXN). Fungal type KRs have a TGXXXGX(1-2)G NAD(P)-binding motif. Some atypical SDRs have lost catalytic activity and/or have an unusual NAD(P)-binding motif and missing or unusual active site residues. Reactions catalyzed within the SDR family include isomerization, decarboxylation, epimerization, C=N bond reduction, dehydratase activity, dehalogenation, Enoyl-CoA reduction, and carbonyl-alcohol oxidoreduction. Pssm-ID: 187659 [Multi-domain] Cd Length: 448 Bit Score: 117.75 E-value: 2.43e-28
|
||||||||||||
| Name | Accession | Description | Interval | E-value | ||||||||
| PS-DH | pfam14765 | Polyketide synthase dehydratase; This is the dehydratase domain of polyketide synthases. ... |
85-377 | 4.59e-52 | ||||||||
Polyketide synthase dehydratase; This is the dehydratase domain of polyketide synthases. Structural analysis shows these DH domains are double hotdogs in which the active site contains a histidine from the N-terminal hotdog and an aspartate from the C-terminal hotdog. Studies have uncovered that a substrate tunnel formed between the DH domains may be essential for loading substrates and unloading products. Pssm-ID: 434191 Cd Length: 296 Bit Score: 179.49 E-value: 4.59e-52
|
||||||||||||
| PKS_DH | smart00826 | Dehydratase domain in polyketide synthase (PKS) enzymes; |
85-259 | 2.56e-49 | ||||||||
Dehydratase domain in polyketide synthase (PKS) enzymes; Pssm-ID: 214837 Cd Length: 167 Bit Score: 167.79 E-value: 2.56e-49
|
||||||||||||
| PksD | COG3321 | Acyl transferase domain in polyketide synthase (PKS) enzymes [Secondary metabolites ... |
2-543 | 1.41e-29 | ||||||||
Acyl transferase domain in polyketide synthase (PKS) enzymes [Secondary metabolites biosynthesis, transport and catabolism]; Pssm-ID: 442550 [Multi-domain] Cd Length: 1386 Bit Score: 124.21 E-value: 1.41e-29
|
||||||||||||
| KR_3_FAS_SDR_x | cd08956 | beta-ketoacyl reductase (KR) domain of fatty acid synthase (FAS), subgroup 3, complex (x); ... |
384-543 | 2.43e-28 | ||||||||
beta-ketoacyl reductase (KR) domain of fatty acid synthase (FAS), subgroup 3, complex (x); Ketoreductase, a module of the multidomain polyketide synthase (PKS), has 2 subdomains, each corresponding to a SDR family monomer. The C-terminal subdomain catalyzes the NADPH-dependent reduction of the beta-carbonyl of a polyketide to a hydroxyl group, a step in the biosynthesis of polyketides, such as erythromycin. The N-terminal subdomain, an interdomain linker, is a truncated Rossmann fold which acts to stabilizes the catalytic subdomain. Unlike typical SDRs, the isolated domain does not oligomerize but is composed of 2 subdomains, each resembling an SDR monomer. The active site resembles that of typical SDRs, except that the usual positions of the catalytic Asn and Tyr are swapped, so that the canonical YXXXK motif changes to YXXXN. Modular PKSs are multifunctional structures in which the makeup recapitulates that found in (and may have evolved from) FAS. In some instances, such as porcine FAS, an enoyl reductase (ER) module is inserted between the sub-domains. Fatty acid synthesis occurs via the stepwise elongation of a chain (which is attached to acyl carrier protein, ACP) with 2-carbon units. Eukaryotic systems consists of large, multifunctional synthases (type I) while bacterial, type II systems, use single function proteins. Fungal fatty acid synthesis uses a dodecamer of 6 alpha and 6 beta subunits. In mammalian type FAS cycles, ketoacyl synthase forms acetoacetyl-ACP which is reduced by the NADP-dependent beta-KR, forming beta-hydroxyacyl-ACP, which is in turn dehydrated by dehydratase to a beta-enoyl intermediate, which is reduced by NADP-dependent beta- ER. Polyketide synthesis also proceeds via the addition of 2-carbon units as in fatty acid synthesis. The complex SDR NADP-binding motif, GGXGXXG, is often present, but is not strictly conserved in each instance of the module. This subfamily includes KR domains found in many multidomain PKSs, including six of seven Sorangium cellulosum PKSs (encoded by spiDEFGHIJ) which participate in the synthesis of the polyketide scaffold of the cytotoxic spiroketal polyketide spirangien. These seven PKSs have either a single PKS module (SpiF), two PKR modules (SpiD,-E,-I,-J), or three PKS modules (SpiG,-H). This subfamily includes the second KR domains of SpiE,-G, I, and -J, both KR domains of SpiD, and the third KR domain of SpiH. The single KR domain of SpiF, the first and second KR domains of SpiH, the first KR domains of SpiE,-G,- I, and -J, and the third KR domain of SpiG, belong to a different KR_FAS_SDR subfamily. SDRs are a functionally diverse family of oxidoreductases that have a single domain with a structurally conserved Rossmann fold (alpha/beta folding pattern with a central beta-sheet), an NAD(P)(H)-binding region, and a structurally diverse C-terminal region. Classical SDRs are typically about 250 residues long, while extended SDRs are approximately 350 residues. Sequence identity between different SDR enzymes are typically in the 15-30% range, but the enzymes share the Rossmann fold NAD-binding motif and characteristic NAD-binding and catalytic sequence patterns. These enzymes catalyze a wide range of activities including the metabolism of steroids, cofactors, carbohydrates, lipids, aromatic compounds, and amino acids, and act in redox sensing. Classical SDRs have an TGXXX[AG]XG cofactor binding motif and a YXXXK active site motif, with the Tyr residue of the active site motif serving as a critical catalytic residue (Tyr-151, human prostaglandin dehydrogenase (PGDH) numbering). In addition to the Tyr and Lys, there is often an upstream Ser (Ser-138, PGDH numbering) and/or an Asn (Asn-107, PGDH numbering) contributing to the active site; while substrate binding is in the C-terminal region, which determines specificity. The standard reaction mechanism is a 4-pro-S hydride transfer and proton relay involving the conserved Tyr and Lys, a water molecule stabilized by Asn, and nicotinamide. Extended SDRs have additional elements in the C-terminal region, and typically have a TGXXGXXG cofactor binding motif. Complex (multidomain) SDRs such as ketoreductase domains of fatty acid synthase have a GGXGXXG NAD(P)-binding motif and an altered active site motif (YXXXN). Fungal type KRs have a TGXXXGX(1-2)G NAD(P)-binding motif. Some atypical SDRs have lost catalytic activity and/or have an unusual NAD(P)-binding motif and missing or unusual active site residues. Reactions catalyzed within the SDR family include isomerization, decarboxylation, epimerization, C=N bond reduction, dehydratase activity, dehalogenation, Enoyl-CoA reduction, and carbonyl-alcohol oxidoreduction. Pssm-ID: 187659 [Multi-domain] Cd Length: 448 Bit Score: 117.75 E-value: 2.43e-28
|
||||||||||||
| KR_2_FAS_SDR_x | cd08955 | beta-ketoacyl reductase (KR) domain of fatty acid synthase (FAS), subgroup 2, complex (x); ... |
437-543 | 1.08e-06 | ||||||||
beta-ketoacyl reductase (KR) domain of fatty acid synthase (FAS), subgroup 2, complex (x); Ketoreductase, a module of the multidomain polyketide synthase, has 2 subdomains, each corresponding to a short-chain dehydrogenases/reductase (SDR) family monomer. The C-terminal subdomain catalyzes the NADPH-dependent reduction of the beta-carbonyl of a polyketide to a hydroxyl group, a step in the biosynthesis of polyketides, such as erythromycin. The N-terminal subdomain, an interdomain linker, is a truncated Rossmann fold which acts to stabilizes the catalytic subdomain. Unlike typical SDRs, the isolated domain does not oligomerizes but is composed of 2 subdomains, each resembling an SDR monomer. In some instances, as in porcine FAS, an enoyl reductase (a Rossman fold NAD binding domain of the MDR family) module is inserted between the sub-domains. The active site resembles that of typical SDRs, except that the usual positions of the catalytic asparagine and tyrosine are swapped, so that the canonical YXXXK motif changes to YXXXN. Modular polyketide synthases are multifunctional structures in which the makeup recapitulates that found in (and may have evolved from) fatty acid synthase. In some instances, such as porcine FAS , an enoyl reductase module is inserted between the sub-domains. Fatty acid synthesis occurs via the stepwise elongation of a chain (which is attached to acyl carrier protein, ACP) with 2-carbon units. Eukaryotic systems consists of large, multifunctional synthases (type I) while bacterial, type II systems, use single function proteins. Fungal fatty acid synthesis uses dodecamer of 6 alpha and 6 beta subunits. In mammalian type FAS cycles, ketoacyl synthase forms acetoacetyl-ACP which is reduced by the NADP-dependent beta-ketoacyl reductase (KR), forming beta-hydroxyacyl-ACP, which is in turn dehydrated by dehydratase to a beta-enoyl intermediate, which is reduced by NADP-dependent beta-enoyl reductase (ER). Polyketide syntheses also proceeds via the addition of 2-carbon units as in fatty acid synthesis. The complex SDR NADP binding motif, GGXGXXG, is often present, but is not strictly conserved in each instance of the module. This subfamily includes the KR domain of the Lyngbya majuscule Jam J, -K, and #L which are encoded on the jam gene cluster and are involved in the synthesis of the Jamaicamides (neurotoxins); Lyngbya majuscule Jam P belongs to a different KR_FAS_SDR_x subfamily. SDRs are a functionally diverse family of oxidoreductases that have a single domain with a structurally conserved Rossmann fold (alpha/beta folding pattern with a central beta-sheet), an NAD(P)(H)-binding region, and a structurally diverse C-terminal region. Classical SDRs are typically about 250 residues long, while extended SDRs are approximately 350 residues. Sequence identity between different SDR enzymes are typically in the 15-30% range, but the enzymes share the Rossmann fold NAD-binding motif and characteristic NAD-binding and catalytic sequence patterns. These enzymes catalyze a wide range of activities including the metabolism of steroids, cofactors, carbohydrates, lipids, aromatic compounds, and amino acids, and act in redox sensing. Classical SDRs have an TGXXX[AG]XG cofactor binding motif and a YXXXK active site motif, with the Tyr residue of the active site motif serving as a critical catalytic residue (Tyr-151, human prostaglandin dehydrogenase (PGDH) numbering). In addition to the Tyr and Lys, there is often an upstream Ser (Ser-138, PGDH numbering) and/or an Asn (Asn-107, PGDH numbering) contributing to the active site; while substrate binding is in the C-terminal region, which determines specificity. The standard reaction mechanism is a 4-pro-S hydride transfer and proton relay involving the conserved Tyr and Lys, a water molecule stabilized by Asn, and nicotinamide. Extended SDRs have additional elements in the C-terminal region, and typically have a TGXXGXXG cofactor binding motif. Complex (multidomain) SDRs such as ketoreductase domains of fatty acid synthase have a GGXGXXG NAD(P)-binding motif and an altered active site motif (YXXXN). Fungal type KRs have a TGXXXGX(1-2)G NAD(P)-binding motif. Some atypical SDRs have lost catalytic activity and/or have an unusual NAD(P)-binding motif and missing or unusual active site residues. Reactions catalyzed within the SDR family include isomerization, decarboxylation, epimerization, C=N bond reduction, dehydratase activity, dehalogenation, Enoyl-CoA reduction, and carbonyl-alcohol oxidoreduction. Pssm-ID: 187658 [Multi-domain] Cd Length: 376 Bit Score: 50.75 E-value: 1.08e-06
|
||||||||||||
| KR_FAS_SDR_x | cd05274 | ketoreductase (KR) and fatty acid synthase (FAS), complex (x) SDRs; Ketoreductase, a module of ... |
436-543 | 7.59e-05 | ||||||||
ketoreductase (KR) and fatty acid synthase (FAS), complex (x) SDRs; Ketoreductase, a module of the multidomain polyketide synthase (PKS), has 2 subdomains, each corresponding to a SDR family monomer. The C-terminal subdomain catalyzes the NADPH-dependent reduction of the beta-carbonyl of a polyketide to a hydroxyl group, a step in the biosynthesis of polyketides, such as erythromycin. The N-terminal subdomain, an interdomain linker, is a truncated Rossmann fold which acts to stabilizes the catalytic subdomain. Unlike typical SDRs, the isolated domain does not oligomerize but is composed of 2 subdomains, each resembling an SDR monomer. The active site resembles that of typical SDRs, except that the usual positions of the catalytic Asn and Tyr are swapped, so that the canonical YXXXK motif changes to YXXXN. Modular PKSs are multifunctional structures in which the makeup recapitulates that found in (and may have evolved from) FAS. In some instances, such as porcine FAS, an enoyl reductase (ER) module is inserted between the sub-domains. Fatty acid synthesis occurs via the stepwise elongation of a chain (which is attached to acyl carrier protein, ACP) with 2-carbon units. Eukaryotic systems consist of large, multifunctional synthases (type I) while bacterial, type II systems, use single function proteins. Fungal fatty acid synthase uses a dodecamer of 6 alpha and 6 beta subunits. In mammalian type FAS cycles, ketoacyl synthase forms acetoacetyl-ACP which is reduced by the NADP-dependent beta-KR, forming beta-hydroxyacyl-ACP, which is in turn dehydrated by dehydratase to a beta-enoyl intermediate, which is reduced by NADP-dependent beta-ER. Polyketide synthesis also proceeds via the addition of 2-carbon units as in fatty acid synthesis. The complex SDR NADP-binding motif, GGXGXXG, is often present, but is not strictly conserved in each instance of the module. SDRs are a functionally diverse family of oxidoreductases that have a single domain with a structurally conserved Rossmann fold (alpha/beta folding pattern with a central beta-sheet), an NAD(P)(H)-binding region, and a structurally diverse C-terminal region. Classical SDRs are typically about 250 residues long, while extended SDRs are approximately 350 residues. Sequence identity between different SDR enzymes are typically in the 15-30% range, but the enzymes share the Rossmann fold NAD-binding motif and characteristic NAD-binding and catalytic sequence patterns. These enzymes catalyze a wide range of activities including the metabolism of steroids, cofactors, carbohydrates, lipids, aromatic compounds, and amino acids, and act in redox sensing. Classical SDRs have an TGXXX[AG]XG cofactor binding motif and a YXXXK active site motif, with the Tyr residue of the active site motif serving as a critical catalytic residue (Tyr-151, human prostaglandin dehydrogenase (PGDH) numbering). In addition to the Tyr and Lys, there is often an upstream Ser (Ser-138, PGDH numbering) and/or an Asn (Asn-107, PGDH numbering) contributing to the active site; while substrate binding is in the C-terminal region, which determines specificity. The standard reaction mechanism is a 4-pro-S hydride transfer and proton relay involving the conserved Tyr and Lys, a water molecule stabilized by Asn, and nicotinamide. Extended SDRs have additional elements in the C-terminal region, and typically have a TGXXGXXG cofactor binding motif. Complex (multidomain) SDRs such as ketoreductase domains of fatty acid synthase have a GGXGXXG NAD(P)-binding motif and an altered active site motif (YXXXN). Fungal type KRs have a TGXXXGX(1-2)G NAD(P)-binding motif. Some atypical SDRs have lost catalytic activity and/or have an unusual NAD(P)-binding motif and missing or unusual active site residues. Reactions catalyzed within the SDR family include isomerization, decarboxylation, epimerization, C=N bond reduction, dehydratase activity, dehalogenation, Enoyl-CoA reduction, and carbonyl-alcohol oxidoreduction. Pssm-ID: 187582 [Multi-domain] Cd Length: 375 Bit Score: 45.07 E-value: 7.59e-05
|
||||||||||||
| KR_1_SDR_x | cd08952 | ketoreductase (KR), subgroup 1, complex (x) SDRs; Ketoreductase, a module of the multidomain ... |
471-543 | 3.20e-03 | ||||||||
ketoreductase (KR), subgroup 1, complex (x) SDRs; Ketoreductase, a module of the multidomain polyketide synthase (PKS), has 2 subdomains, each corresponding to a SDR family monomer. The C-terminal subdomain catalyzes the NADPH-dependent reduction of the beta-carbonyl of a polyketide to a hydroxyl group, a step in the biosynthesis of polyketides, such as erythromycin. The N-terminal subdomain, an interdomain linker, is a truncated Rossmann fold which acts to stabilizes the catalytic subdomain. Unlike typical SDRs, the isolated domain does not oligomerize but is composed of 2 subdomains, each resembling an SDR monomer. The active site resembles that of typical SDRs, except that the usual positions of the catalytic Asn and Tyr are swapped, so that the canonical YXXXK motif changes to YXXXN. Modular PKSs are multifunctional structures in which the makeup recapitulates that found in (and may have evolved from) FAS. Polyketide synthesis also proceeds via the addition of 2-carbon units as in fatty acid synthesis. The complex SDR NADP-binding motif, GGXGXXG, is often present, but is not strictly conserved in each instance of the module. This subfamily includes KR domains found in many multidomain PKSs, including six of seven Sorangium cellulosum PKSs (encoded by spiDEFGHIJ) which participate in the synthesis of the polyketide scaffold of the cytotoxic spiroketal polyketide spirangien. These seven PKSs have either a single PKS module (SpiF), two PKR modules (SpiD,-E,-I,-J), or three PKS modules (SpiG,-H). This subfamily includes the single KR domain of SpiF, the first KR domains of SpiE,-G,H,-I,and #J, the third KR domain of SpiG, and the second KR domain of SpiH. The second KR domains of SpiE,-G, I, and #J, and the KR domains of SpiD, belong to a different KR_FAS_SDR subfamily. SDRs are a functionally diverse family of oxidoreductases that have a single domain with a structurally conserved Rossmann fold (alpha/beta folding pattern with a central beta-sheet), an NAD(P)(H)-binding region, and a structurally diverse C-terminal region. Classical SDRs are typically about 250 residues long, while extended SDRs are approximately 350 residues. Sequence identity between different SDR enzymes are typically in the 15-30% range, but the enzymes share the Rossmann fold NAD-binding motif and characteristic NAD-binding and catalytic sequence patterns. These enzymes catalyze a wide range of activities including the metabolism of steroids, cofactors, carbohydrates, lipids, aromatic compounds, and amino acids, and act in redox sensing. Classical SDRs have an TGXXX[AG]XG cofactor binding motif and a YXXXK active site motif, with the Tyr residue of the active site motif serving as a critical catalytic residue (Tyr-151, human prostaglandin dehydrogenase (PGDH) numbering). In addition to the Tyr and Lys, there is often an upstream Ser (Ser-138, PGDH numbering) and/or an Asn (Asn-107, PGDH numbering) contributing to the active site; while substrate binding is in the C-terminal region, which determines specificity. The standard reaction mechanism is a 4-pro-S hydride transfer and proton relay involving the conserved Tyr and Lys, a water molecule stabilized by Asn, and nicotinamide. Extended SDRs have additional elements in the C-terminal region, and typically have a TGXXGXXG cofactor binding motif. Complex (multidomain) SDRs such as ketoreductase domains of fatty acid synthase have a GGXGXXG NAD(P)-binding motif and an altered active site motif (YXXXN). Fungal type KRs have a TGXXXGX(1-2)G NAD(P)-binding motif. Some atypical SDRs have lost catalytic activity and/or have an unusual NAD(P)-binding motif and missing or unusual active site residues. Reactions catalyzed within the SDR family include isomerization, decarboxylation, epimerization, C=N bond reduction, dehydratase activity, dehalogenation, Enoyl-CoA reduction, and carbonyl-alcohol oxidoreduction. Pssm-ID: 187655 [Multi-domain] Cd Length: 480 Bit Score: 40.23 E-value: 3.20e-03
|
||||||||||||
| Blast search parameters | ||||
|
