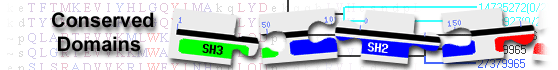glycoside hydrolase family 32 protein similar to invertase, which hydrolyzes terminal non-reducing beta-D-fructofuranoside residues in beta-D-fructofuranosides
glycoside hydrolase family 32 sucrose 6 phosphate hydrolase (sucrase); Glycosyl hydrolase family GH32 subgroup contains sucrose-6-phosphate hydrolase (sucrase, EC:3.2.1.26) among others. The enzyme cleaves sucrose into fructose and glucose via beta-fructofuranosidase activity, producing invert sugar that is a mixture of dextrorotatory D-glucose and levorotatory D-fructose. These retaining enzymes (i.e. they retain the configuration at anomeric carbon atom of the substrate) catalyze hydrolysis in two steps involving a covalent glycosyl enzyme intermediate: an aspartate located close to the N-terminus acts as the catalytic nucleophile and a glutamate acts as the general acid/base; a conserved aspartate residue in the Arg-Asp-Pro (RDP) motif stabilizes the transition state. The breakdown of sucrose is widely used as a carbon or energy source by bacteria, fungi, and plants. Invertase is used commercially in the confectionery industry, since fructose has a sweeter taste than sucrose and a lower tendency to crystallize. A common structural feature of all these enzymes is a 5-bladed beta-propeller domain, similar to GH43, that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller.
:
Pssm-ID: 350135 Cd Length: 289 Bit Score: 391.87 E-value: 5.31e-135
Glycosyl hydrolases family 32 C terminal; This domain corresponds to the C terminal domain of ...
346-493
7.18e-10
Glycosyl hydrolases family 32 C terminal; This domain corresponds to the C terminal domain of glycosyl hydrolase family 32. It forms a beta sandwich module.
The actual alignment was detected with superfamily member pfam08244:
Pssm-ID: 462408 [Multi-domain] Cd Length: 162 Bit Score: 57.75 E-value: 7.18e-10
glycoside hydrolase family 32 sucrose 6 phosphate hydrolase (sucrase); Glycosyl hydrolase family GH32 subgroup contains sucrose-6-phosphate hydrolase (sucrase, EC:3.2.1.26) among others. The enzyme cleaves sucrose into fructose and glucose via beta-fructofuranosidase activity, producing invert sugar that is a mixture of dextrorotatory D-glucose and levorotatory D-fructose. These retaining enzymes (i.e. they retain the configuration at anomeric carbon atom of the substrate) catalyze hydrolysis in two steps involving a covalent glycosyl enzyme intermediate: an aspartate located close to the N-terminus acts as the catalytic nucleophile and a glutamate acts as the general acid/base; a conserved aspartate residue in the Arg-Asp-Pro (RDP) motif stabilizes the transition state. The breakdown of sucrose is widely used as a carbon or energy source by bacteria, fungi, and plants. Invertase is used commercially in the confectionery industry, since fructose has a sweeter taste than sucrose and a lower tendency to crystallize. A common structural feature of all these enzymes is a 5-bladed beta-propeller domain, similar to GH43, that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller.
Pssm-ID: 350135 Cd Length: 289 Bit Score: 391.87 E-value: 5.31e-135
Glycosyl hydrolases family 32 N-terminal domain; This domain corresponds to the N-terminal ...
18-343
6.51e-85
Glycosyl hydrolases family 32 N-terminal domain; This domain corresponds to the N-terminal domain of glycosyl hydrolase family 32 which forms a five bladed beta propeller structure.
Pssm-ID: 425557 Cd Length: 308 Bit Score: 264.50 E-value: 6.51e-85
Glycosyl hydrolases family 32 C terminal; This domain corresponds to the C terminal domain of ...
346-493
7.18e-10
Glycosyl hydrolases family 32 C terminal; This domain corresponds to the C terminal domain of glycosyl hydrolase family 32. It forms a beta sandwich module.
Pssm-ID: 462408 [Multi-domain] Cd Length: 162 Bit Score: 57.75 E-value: 7.18e-10
glycoside hydrolase family 32 sucrose 6 phosphate hydrolase (sucrase); Glycosyl hydrolase family GH32 subgroup contains sucrose-6-phosphate hydrolase (sucrase, EC:3.2.1.26) among others. The enzyme cleaves sucrose into fructose and glucose via beta-fructofuranosidase activity, producing invert sugar that is a mixture of dextrorotatory D-glucose and levorotatory D-fructose. These retaining enzymes (i.e. they retain the configuration at anomeric carbon atom of the substrate) catalyze hydrolysis in two steps involving a covalent glycosyl enzyme intermediate: an aspartate located close to the N-terminus acts as the catalytic nucleophile and a glutamate acts as the general acid/base; a conserved aspartate residue in the Arg-Asp-Pro (RDP) motif stabilizes the transition state. The breakdown of sucrose is widely used as a carbon or energy source by bacteria, fungi, and plants. Invertase is used commercially in the confectionery industry, since fructose has a sweeter taste than sucrose and a lower tendency to crystallize. A common structural feature of all these enzymes is a 5-bladed beta-propeller domain, similar to GH43, that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller.
Pssm-ID: 350135 Cd Length: 289 Bit Score: 391.87 E-value: 5.31e-135
Glycosyl hydrolases family 32 N-terminal domain; This domain corresponds to the N-terminal ...
18-343
6.51e-85
Glycosyl hydrolases family 32 N-terminal domain; This domain corresponds to the N-terminal domain of glycosyl hydrolase family 32 which forms a five bladed beta propeller structure.
Pssm-ID: 425557 Cd Length: 308 Bit Score: 264.50 E-value: 6.51e-85
Glycosyl hydrolase family 32, beta-fructosidases; Glycosyl hydrolase family GH32 cleaves ...
28-333
2.41e-72
Glycosyl hydrolase family 32, beta-fructosidases; Glycosyl hydrolase family GH32 cleaves sucrose into fructose and glucose via beta-fructofuranosidase activity, producing invert sugar that is a mixture of dextrorotatory D-glucose and levorotatory D-fructose, thus named invertase (EC 3.2.1.26). This family also contains other fructofuranosidases such as inulinase (EC 3.2.1.7), exo-inulinase (EC 3.2.1.80), levanase (EC 3.2.1.65), and transfructosidases such sucrose:sucrose 1-fructosyltransferase (EC 2.4.1.99), fructan:fructan 1-fructosyltransferase (EC 2.4.1.100), sucrose:fructan 6-fructosyltransferase (EC 2.4.1.10), fructan:fructan 6G-fructosyltransferase (EC 2.4.1.243) and levan fructosyltransferases (EC 2.4.1.-). These retaining enzymes (i.e. they retain the configuration at anomeric carbon atom of the substrate) catalyze hydrolysis in two steps involving a covalent glycosyl enzyme intermediate: an aspartate located close to the N-terminus acts as the catalytic nucleophile and a glutamate acts as the general acid/base; a conserved aspartate residue in the Arg-Asp-Pro (RDP) motif stabilizes the transition state. These enzymes are predicted to display a 5-fold beta-propeller fold as found for GH43 and CH68. The breakdown of sucrose is widely used as a carbon or energy source by bacteria, fungi, and plants. Invertase is used commercially in the confectionery industry, since fructose has a sweeter taste than sucrose and a lower tendency to crystallize. A common structural feature of all these enzymes is a 5-bladed beta-propeller domain, similar to GH43, that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller.
Pssm-ID: 350110 Cd Length: 281 Bit Score: 230.99 E-value: 2.41e-72
glycoside hydrolase family 32 protein such as Thermotoga maritima invertase (BfrA or Tm1414); ...
28-333
3.67e-47
glycoside hydrolase family 32 protein such as Thermotoga maritima invertase (BfrA or Tm1414); This subfamily of glycosyl hydrolase family GH32 includes beta-fructosidase (invertase, EC 3.2.1.26) that cleaves sucrose into fructose and glucose via beta-fructofuranosidase activity, producing invert sugar that is a mixture of dextrorotatory D-glucose and levorotatory D-fructose, thus named invertase. These retaining enzymes (i.e. they retain the configuration at anomeric carbon atom of the substrate) catalyze hydrolysis in two steps involving a covalent glycosyl enzyme intermediate: an aspartate located close to the N-terminus acts as the catalytic nucleophile and a glutamate acts as the general acid/base; a conserved aspartate residue in the Arg-Asp-Pro (RDP) motif stabilizes the transition state. The breakdown of sucrose is widely used as a carbon or energy source by bacteria, fungi, and plants. Invertase is used commercially in the confectionery industry, since fructose has a sweeter taste than sucrose and a lower tendency to crystallize. A common structural feature of all these enzymes is a 5-bladed beta-propeller domain, similar to GH43, that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller.
Pssm-ID: 350137 Cd Length: 286 Bit Score: 165.15 E-value: 3.67e-47
glycoside hydrolase family 32 protein such as Aspergillus ficuum endo-inulinase (Inu2); This ...
28-333
1.13e-31
glycoside hydrolase family 32 protein such as Aspergillus ficuum endo-inulinase (Inu2); This subfamily of glycosyl hydrolase family GH32 includes endo-inulinase (inu2, EC 3.2.1.7), exo-inulinase (Inu1, EC 3.2.1.80), invertase (EC 3.2.1.26), and levan fructotransferase (LftA, EC 4.2.2.16), among others. These enzymes cleave sucrose into fructose and glucose via beta-fructofuranosidase activity, producing invert sugar that is a mixture of dextrorotatory D-glucose and levorotatory D-fructose, thus named invertase (EC 3.2.1.26). These retaining enzymes (i.e. they retain the configuration at anomeric carbon atom of the substrate) catalyze hydrolysis in two steps involving a covalent glycosyl enzyme intermediate: an aspartate located close to the N-terminus acts as the catalytic nucleophile and a glutamate acts as the general acid/base; a conserved aspartate residue in the Arg-Asp-Pro (RDP) motif stabilizes the transition state. These enzymes are predicted to display a 5-fold beta-propeller fold as found for GH43 and CH68. The breakdown of sucrose is widely used as a carbon or energy source by bacteria, fungi, and plants. Invertase is used commercially in the confectionery industry, since fructose has a sweeter taste than sucrose and a lower tendency to crystallize. A common structural feature of all these enzymes is a 5-bladed beta-propeller domain, similar to GH43, that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller.
Pssm-ID: 350134 Cd Length: 289 Bit Score: 123.49 E-value: 1.13e-31
Glycosyl hydrolase family 32, such as the putative glycoside hydrolase Escherichia coli Aec43 ...
36-223
2.54e-27
Glycosyl hydrolase family 32, such as the putative glycoside hydrolase Escherichia coli Aec43 (FosGH2); This glycosyl hydrolase family 32 (GH32) subgroup includes Escherichia coli strain BEN2908 putative glycoside hydrolase Aec43 (FosGH2). GH32 enzymes cleave sucrose into fructose and glucose via beta-fructofuranosidase activity, producing invert sugar that is a mixture of dextrorotatory D-glucose and levorotatory D-fructose, thus named invertase (EC 3.2.1.26). GH32 family also contains other fructofuranosidases such as inulinase (EC 3.2.1.7), exo-inulinase (EC 3.2.1.80), levanase (EC 3.2.1.65), and transfructosidases such sucrose:sucrose 1-fructosyltransferase (EC 2.4.1.99), fructan:fructan 1-fructosyltransferase (EC 2.4.1.100), sucrose:fructan 6-fructosyltransferase (EC 2.4.1.10), fructan:fructan 6G-fructosyltransferase (EC 2.4.1.243) and levan fructosyltransferases (EC 2.4.1.-). These retaining enzymes (i.e. they retain the configuration at anomeric carbon atom of the substrate) catalyze hydrolysis in two steps involving a covalent glycosyl enzyme intermediate: an aspartate located close to the N-terminus acts as the catalytic nucleophile and a glutamate acts as the general acid/base; a conserved aspartate residue in the Arg-Asp-Pro (RDP) motif stabilizes the transition state. These enzymes are predicted to display a 5-fold beta-propeller fold as found for GH43 and CH68. The breakdown of sucrose is widely used as a carbon or energy source by bacteria, fungi, and plants. Invertase is used commercially in the confectionery industry, since fructose has a sweeter taste than sucrose and a lower tendency to crystallize.
Pssm-ID: 350109 [Multi-domain] Cd Length: 281 Bit Score: 111.13 E-value: 2.54e-27
glycoside hydrolase family 32 protein such as Arabidopsis thaliana cell-wall invertase 1 ...
28-333
3.37e-22
glycoside hydrolase family 32 protein such as Arabidopsis thaliana cell-wall invertase 1 (AtBFruct1;Fruct1;AtcwINV1;At3g13790); This subfamily of glycosyl hydrolase family GH32 includes fructan beta-(2,1)-fructosidase and fructan 1-exohydrolase IIa (1-FEH IIa, EC 3.2.1.153), cell-wall invertase 1 (EC 3.2.1.26), sucrose:fructan 6-fructosyltransferase (6-Sst/6-Dft, EC 2.4.1.10), and levan fructosyltransferases (EC 2.4.1.-) among others. This enzyme cleaves sucrose into fructose and glucose via beta-fructofuranosidase activity, producing invert sugar that is a mixture of dextrorotatory D-glucose and levorotatory D-fructose, thus named invertase. These retaining enzymes (i.e. they retain the configuration at anomeric carbon atom of the substrate) catalyze hydrolysis in two steps involving a covalent glycosyl enzyme intermediate: an aspartate located close to the N-terminus acts as the catalytic nucleophile and a glutamate acts as the general acid/base; a conserved aspartate residue in the Arg-Asp-Pro (RDP) motif stabilizes the transition state. The breakdown of sucrose is widely used as a carbon or energy source by bacteria, fungi, and plants. Invertase is used commercially in the confectionery industry, since fructose has a sweeter taste than sucrose and a lower tendency to crystallize. A common structural feature of all these enzymes is a 5-bladed beta-propeller domain, similar to GH43, that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller.
Pssm-ID: 350136 [Multi-domain] Cd Length: 296 Bit Score: 96.69 E-value: 3.37e-22
Glycosyl hydrolase families 32 and 68, which form the clan GH-J; This glycosyl hydrolase ...
28-331
1.75e-16
Glycosyl hydrolase families 32 and 68, which form the clan GH-J; This glycosyl hydrolase family clan J (according to carbohydrate-active enzymes database (CAZY)) includes family 32 (GH32) and 68 (GH68). GH32 enzymes include invertase (EC 3.2.1.26) and other other fructofuranosidases such as inulinase (EC 3.2.1.7), exo-inulinase (EC 3.2.1.80), levanase (EC 3.2.1.65), and transfructosidases such sucrose:sucrose 1-fructosyltransferase (EC 2.4.1.99), fructan:fructan 1-fructosyltransferase (EC 2.4.1.100), sucrose:fructan 6-fructosyltransferase (EC 2.4.1.10), fructan:fructan 6G-fructosyltransferase (EC 2.4.1.243) and levan fructosyltransferases (EC 2.4.1.-). The GH68 family consists of frucosyltransferases (FTFs) that include levansucrase (EC 2.4.1.10, also known as beta-D-fructofuranosyl transferase), beta-fructofuranosidase (EC 3.2.1.26) and inulosucrase (EC 2.4.1.9). GH32 and GH68 family enzymes are retaining enzymes (i.e. they retain the configuration at anomeric carbon atom of the substrate) and catalyze hydrolysis in two steps involving a covalent glycosyl enzyme intermediate: an aspartate located close to the N-terminus acts as the catalytic nucleophile and a glutamate acts as the general acid/base; a conserved aspartate residue in the Arg-Asp-Pro (RDP) motif stabilizes the transition state. A common structural feature of all these enzymes is a 5-bladed beta-propeller domain, similar to GH43, that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller.
Pssm-ID: 350093 [Multi-domain] Cd Length: 292 Bit Score: 79.92 E-value: 1.75e-16
Glycosyl hydrolase family 32 family protein; The GH32 family contains glycosyl hydrolase ...
36-304
4.77e-14
Glycosyl hydrolase family 32 family protein; The GH32 family contains glycosyl hydrolase family GH32 proteins that cleave sucrose into fructose and glucose via beta-fructofuranosidase activity, producing invert sugar that is a mixture of dextrorotatory D-glucose and levorotatory D-fructose, thus named invertase (EC 3.2.1.26). This family also contains other fructofuranosidases such as inulinase (EC 3.2.1.7), exo-inulinase (EC 3.2.1.80), levanase (EC 3.2.1.65), and transfructosidases such sucrose:sucrose 1-fructosyltransferase (EC 2.4.1.99), fructan:fructan 1-fructosyltransferase (EC 2.4.1.100), sucrose:fructan 6-fructosyltransferase (EC 2.4.1.10), fructan:fructan 6G-fructosyltransferase (EC 2.4.1.243) and levan fructosyltransferases (EC 2.4.1.-). These retaining enzymes (i.e. they retain the configuration at anomeric carbon atom of the substrate) catalyze hydrolysis in two steps involving a covalent glycosyl enzyme intermediate: an aspartate located close to the N-terminus acts as the catalytic nucleophile and a glutamate acts as the general acid/base; a conserved aspartate residue in the Arg-Asp-Pro (RDP) motif stabilizes the transition state. These enzymes are predicted to display a 5-fold beta-propeller fold as found for GH43 and CH68. The breakdown of sucrose is widely used as a carbon or energy source by bacteria, fungi, and plants. Invertase is used commercially in the confectionery industry, since fructose has a sweeter taste than sucrose and a lower tendency to crystallize. A common structural feature of all these enzymes is a 5-bladed beta-propeller domain, similar to GH43, that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller.
Pssm-ID: 350121 Cd Length: 303 Bit Score: 72.67 E-value: 4.77e-14
glycoside hydrolase family 32 protein such as Xanthophyllomyces dendrorhous ...
28-333
8.11e-14
glycoside hydrolase family 32 protein such as Xanthophyllomyces dendrorhous beta-fructofuranosidase (Inv;Xd-INV;XdINV); This subfamily of glycosyl hydrolase family GH32 includes fructan:fructan 1-fructosyltransferase (FT, EC 2.4.1.100) and beta-fructofuranosidase (invertase or Inv, EC 3.2.1.26), among others. These enzymes cleave sucrose into fructose and glucose via beta-fructofuranosidase activity, producing invert sugar that is a mixture of dextrorotatory D-glucose and levorotatory D-fructose, thus named invertase (EC 3.2.1.26). These retaining enzymes (i.e. they retain the configuration at anomeric carbon atom of the substrate) catalyze hydrolysis in two steps involving a covalent glycosyl enzyme intermediate: an aspartate located close to the N-terminus acts as the catalytic nucleophile and a glutamate acts as the general acid/base; a conserved aspartate residue in the Arg-Asp-Pro (RDP) motif stabilizes the transition state. Xanthophyllomyces dendrorhous beta-fructofuranosidase (XdINV) also catalyzes the synthesis of fructooligosaccharides (FOS, a beneficial prebiotic), producing neo-FOS, making it an interesting biotechnology target. Structural studies show plasticity of its active site, having a flexible loop that is essential in binding sucrose and beta(2-1)-linked oligosaccharide, making it a valuable biocatalyst to produce novel bioconjugates. The breakdown of sucrose is widely used as a carbon or energy source by bacteria, fungi, and plants. Invertase is used commercially in the confectionery industry, since fructose has a sweeter taste than sucrose and a lower tendency to crystallize. A common structural feature of all these enzymes is a 5-bladed beta-propeller domain, similar to GH43, that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller.
Pssm-ID: 350133 Cd Length: 337 Bit Score: 72.66 E-value: 8.11e-14
Glycosyl hydrolases family 32 C terminal; This domain corresponds to the C terminal domain of ...
346-493
7.18e-10
Glycosyl hydrolases family 32 C terminal; This domain corresponds to the C terminal domain of glycosyl hydrolase family 32. It forms a beta sandwich module.
Pssm-ID: 462408 [Multi-domain] Cd Length: 162 Bit Score: 57.75 E-value: 7.18e-10
Glycosyl hydrolase family 43; This glycosyl hydrolase family 43 (GH43)-like subfamily includes ...
36-224
2.60e-03
Glycosyl hydrolase family 43; This glycosyl hydrolase family 43 (GH43)-like subfamily includes uncharacterized enzymes similar to those with beta-1,4-xylosidase (xylan 1,4-beta-xylosidase; EC 3.2.1.37), beta-1,3-xylosidase (EC 3.2.1.-), alpha-L-arabinofuranosidase (EC 3.2.1.55), arabinanase (EC 3.2.1.99), xylanase (EC 3.2.1.8), endo-alpha-L-arabinanase and galactan 1,3-beta-galactosidase (EC 3.2.1.145) activities. These are inverting enzymes (i.e. they invert the stereochemistry of the anomeric carbon atom of the substrate) that have an aspartate as the catalytic general base, a glutamate as the catalytic general acid and another aspartate that is responsible for pKa modulation and orienting the catalytic acid. Many of the enzymes in this family display both alpha-L-arabinofuranosidase and beta-D-xylosidase activity using aryl-glycosides as substrates. A common structural feature of GH43 enzymes is a 5-bladed beta-propeller domain that contains the catalytic acid and catalytic base. A long V-shaped groove, partially enclosed at one end, forms a single extended substrate-binding surface across the face of the propeller.
Pssm-ID: 350098 Cd Length: 291 Bit Score: 39.92 E-value: 2.60e-03
Database: CDSEARCH/cdd Low complexity filter: no Composition Based Adjustment: yes E-value threshold: 0.01
References:
Wang J et al. (2023), "The conserved domain database in 2023", Nucleic Acids Res.51(D)384-8.
Lu S et al. (2020), "The conserved domain database in 2020", Nucleic Acids Res.48(D)265-8.
Marchler-Bauer A et al. (2017), "CDD/SPARCLE: functional classification of proteins via subfamily domain architectures.", Nucleic Acids Res.45(D)200-3.
of the residues that compose this conserved feature have been mapped to the query sequence.
Click on the triangle to view details about the feature, including a multiple sequence alignment
of your query sequence and the protein sequences used to curate the domain model,
where hash marks (#) above the aligned sequences show the location of the conserved feature residues.
The thumbnail image, if present, provides an approximate view of the feature's location in 3 dimensions.
Click on the triangle for interactive 3D structure viewing options.
Functional characterization of the conserved domain architecture found on the query.
Click here to see more details.
This image shows a graphical summary of conserved domains identified on the query sequence.
The Show Concise/Full Display button at the top of the page can be used to select the desired level of detail: only top scoring hits
(labeled illustration) or all hits
(labeled illustration).
Domains are color coded according to superfamilies
to which they have been assigned. Hits with scores that pass a domain-specific threshold
(specific hits) are drawn in bright colors.
Others (non-specific hits) and
superfamily placeholders are drawn in pastel colors.
if a domain or superfamily has been annotated with functional sites (conserved features),
they are mapped to the query sequence and indicated through sets of triangles
with the same color and shade of the domain or superfamily that provides the annotation. Mouse over the colored bars or triangles to see descriptions of the domains and features.
click on the bars or triangles to view your query sequence embedded in a multiple sequence alignment of the proteins used to develop the corresponding domain model.
The table lists conserved domains identified on the query sequence. Click on the plus sign (+) on the left to display full descriptions, alignments, and scores.
Click on the domain model's accession number to view the multiple sequence alignment of the proteins used to develop the corresponding domain model.
To view your query sequence embedded in that multiple sequence alignment, click on the colored bars in the Graphical Summary portion of the search results page,
or click on the triangles, if present, that represent functional sites (conserved features)
mapped to the query sequence.
Concise Display shows only the best scoring domain model, in each hit category listed below except non-specific hits, for each region on the query sequence.
(labeled illustration) Standard Display shows only the best scoring domain model from each source, in each hit category listed below for each region on the query sequence.
(labeled illustration) Full Display shows all domain models, in each hit category below, that meet or exceed the RPS-BLAST threshold for statistical significance.
(labeled illustration) Four types of hits can be shown, as available,
for each region on the query sequence:
specific hits meet or exceed a domain-specific e-value threshold
(illustrated example)
and represent a very high confidence that the query sequence belongs to the same protein family as the sequences use to create the domain model
non-specific hits
meet or exceed the RPS-BLAST threshold for statistical significance (default E-value cutoff of 0.01, or an E-value selected by user via the
advanced search options)
the domain superfamily to which the specific and non-specific hits belong
multi-domain models that were computationally detected and are likely to contain multiple single domains
Retrieve proteins that contain one or more of the domains present in the query sequence, using the Conserved Domain Architecture Retrieval Tool
(CDART).
Modify your query to search against a different database and/or use advanced search options