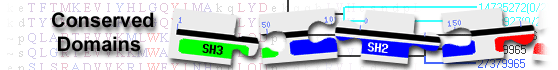cupin domain-containing protein, part of a functionally diverse superfamily with the active site generally located at the center of a conserved domain forming a beta-barrel fold
RmlC-like cupin superfamily; This superfamily contains proteins similar to the RmlC (dTDP ...
3-260
0e+00
RmlC-like cupin superfamily; This superfamily contains proteins similar to the RmlC (dTDP (deoxythymidine diphosphates)-4-dehydrorhamnose 3,5-epimerase)-like cupins. RmlC is a dTDP-sugar isomerase involved in the synthesis of L-rhamnose, a saccharide required for the virulence of some pathogenic bacteria. Cupins are a functionally diverse superfamily originally discovered based on the highly conserved motif found in germin and germin-like proteins. This conserved motif forms a beta-barrel fold found in all of the cupins, giving rise to the name cupin ('cupa' is the Latin term for small barrel). The active site of members of this superfamily is generally located at the center of a conserved barrel and usually includes a metal ion. The different functional classes in this superfamily include single domain bacterial isomerases and epimerases involved in the modification of cell wall carbohydrates, two domain bicupins such as the desiccation-tolerant seed storage globulins, and multidomain nuclear transcription factors involved in legume root nodulation.
The actual alignment was detected with superfamily member TIGR01015:
Pssm-ID: 477354 Cd Length: 429 Bit Score: 511.99 E-value: 0e+00
Homogentisate 1,2-dioxygenase N-terminal; Homogentisate dioxygenase cleaves the aromatic ring ...
5-204
5.59e-127
Homogentisate 1,2-dioxygenase N-terminal; Homogentisate dioxygenase cleaves the aromatic ring during the metabolic degradation of Phe and Tyr. Homogentisate dioxygenase deficiency causes alkaptonuria. The structure of homogentisate dioxygenase shows that the enzyme forms a hexamer arrangement comprised of a dimer of trimers. The active site iron ion is coordinated near the interface between the trimers. This entry represents the N-terminal domain which forms a jelly roll of beta-strands.
Pssm-ID: 466659 Cd Length: 271 Bit Score: 361.28 E-value: 5.59e-127
homogentisate 1,2-dioxygenase and related proteins, N-terminal cupin domain; This family ...
95-183
7.26e-58
homogentisate 1,2-dioxygenase and related proteins, N-terminal cupin domain; This family includes homogentisate 1,2-dioxygenase (also known as homogentisate oxygenase, homogentisic acid oxidase, homogentisicase, HGO, HGD, HGDO, or HmgA; EC 1.13.11.5), which is involved in the metabolic degradation of phenylalanine and tyrosine. It catalyzes the crucial aromatic ring opening reaction, utilizing nonheme Fe2+ to incorporate both atoms of molecular oxygen into homogentisate (2,5-dihydroxyphenylacetate) to yield 4-maleylacetoacetate as part of the homogentisate pathway. HGO deficiency caused by critical mutations and polymorphic sites, causes the metabolic disease alkaptonuria (AKU), a rare disorder of autosomal recessive inheritance. Homogentisate accumulation causes insoluble ochronotic pigments to deposit in connective tissues, resulting in degenerative arthritis. These enzymes are found in prokaryotes, eukaryotes, and archaea. Proteins in this family belong to the cupin superfamily with a conserved "jelly roll-like" beta-barrel fold capable of homodimerization.
Pssm-ID: 380404 [Multi-domain] Cd Length: 109 Bit Score: 180.03 E-value: 7.26e-58
Homogentisate 1,2-dioxygenase N-terminal; Homogentisate dioxygenase cleaves the aromatic ring ...
5-204
5.59e-127
Homogentisate 1,2-dioxygenase N-terminal; Homogentisate dioxygenase cleaves the aromatic ring during the metabolic degradation of Phe and Tyr. Homogentisate dioxygenase deficiency causes alkaptonuria. The structure of homogentisate dioxygenase shows that the enzyme forms a hexamer arrangement comprised of a dimer of trimers. The active site iron ion is coordinated near the interface between the trimers. This entry represents the N-terminal domain which forms a jelly roll of beta-strands.
Pssm-ID: 466659 Cd Length: 271 Bit Score: 361.28 E-value: 5.59e-127
homogentisate 1,2-dioxygenase and related proteins, N-terminal cupin domain; This family ...
95-183
7.26e-58
homogentisate 1,2-dioxygenase and related proteins, N-terminal cupin domain; This family includes homogentisate 1,2-dioxygenase (also known as homogentisate oxygenase, homogentisic acid oxidase, homogentisicase, HGO, HGD, HGDO, or HmgA; EC 1.13.11.5), which is involved in the metabolic degradation of phenylalanine and tyrosine. It catalyzes the crucial aromatic ring opening reaction, utilizing nonheme Fe2+ to incorporate both atoms of molecular oxygen into homogentisate (2,5-dihydroxyphenylacetate) to yield 4-maleylacetoacetate as part of the homogentisate pathway. HGO deficiency caused by critical mutations and polymorphic sites, causes the metabolic disease alkaptonuria (AKU), a rare disorder of autosomal recessive inheritance. Homogentisate accumulation causes insoluble ochronotic pigments to deposit in connective tissues, resulting in degenerative arthritis. These enzymes are found in prokaryotes, eukaryotes, and archaea. Proteins in this family belong to the cupin superfamily with a conserved "jelly roll-like" beta-barrel fold capable of homodimerization.
Pssm-ID: 380404 [Multi-domain] Cd Length: 109 Bit Score: 180.03 E-value: 7.26e-58
Homogentisate 1,2-dioxygenase C-terminal; Homogentisate dioxygenase cleaves the aromatic ring ...
205-260
2.63e-37
Homogentisate 1,2-dioxygenase C-terminal; Homogentisate dioxygenase cleaves the aromatic ring during the metabolic degradation of Phe and Tyr. Homogentisate dioxygenase deficiency causes alkaptonuria. The structure of homogentisate dioxygenase shows that the enzyme forms a hexamer arrangement comprised of a dimer of trimers. The active site iron ion is coordinated near the interface between the trimers. This entry represents the C-terminal active site domain.
Pssm-ID: 461227 [Multi-domain] Cd Length: 153 Bit Score: 128.65 E-value: 2.63e-37
Database: CDSEARCH/cdd Low complexity filter: no Composition Based Adjustment: yes E-value threshold: 0.01
References:
Wang J et al. (2023), "The conserved domain database in 2023", Nucleic Acids Res.51(D)384-8.
Lu S et al. (2020), "The conserved domain database in 2020", Nucleic Acids Res.48(D)265-8.
Marchler-Bauer A et al. (2017), "CDD/SPARCLE: functional classification of proteins via subfamily domain architectures.", Nucleic Acids Res.45(D)200-3.
of the residues that compose this conserved feature have been mapped to the query sequence.
Click on the triangle to view details about the feature, including a multiple sequence alignment
of your query sequence and the protein sequences used to curate the domain model,
where hash marks (#) above the aligned sequences show the location of the conserved feature residues.
The thumbnail image, if present, provides an approximate view of the feature's location in 3 dimensions.
Click on the triangle for interactive 3D structure viewing options.
Functional characterization of the conserved domain architecture found on the query.
Click here to see more details.
This image shows a graphical summary of conserved domains identified on the query sequence.
The Show Concise/Full Display button at the top of the page can be used to select the desired level of detail: only top scoring hits
(labeled illustration) or all hits
(labeled illustration).
Domains are color coded according to superfamilies
to which they have been assigned. Hits with scores that pass a domain-specific threshold
(specific hits) are drawn in bright colors.
Others (non-specific hits) and
superfamily placeholders are drawn in pastel colors.
if a domain or superfamily has been annotated with functional sites (conserved features),
they are mapped to the query sequence and indicated through sets of triangles
with the same color and shade of the domain or superfamily that provides the annotation. Mouse over the colored bars or triangles to see descriptions of the domains and features.
click on the bars or triangles to view your query sequence embedded in a multiple sequence alignment of the proteins used to develop the corresponding domain model.
The table lists conserved domains identified on the query sequence. Click on the plus sign (+) on the left to display full descriptions, alignments, and scores.
Click on the domain model's accession number to view the multiple sequence alignment of the proteins used to develop the corresponding domain model.
To view your query sequence embedded in that multiple sequence alignment, click on the colored bars in the Graphical Summary portion of the search results page,
or click on the triangles, if present, that represent functional sites (conserved features)
mapped to the query sequence.
Concise Display shows only the best scoring domain model, in each hit category listed below except non-specific hits, for each region on the query sequence.
(labeled illustration) Standard Display shows only the best scoring domain model from each source, in each hit category listed below for each region on the query sequence.
(labeled illustration) Full Display shows all domain models, in each hit category below, that meet or exceed the RPS-BLAST threshold for statistical significance.
(labeled illustration) Four types of hits can be shown, as available,
for each region on the query sequence:
specific hits meet or exceed a domain-specific e-value threshold
(illustrated example)
and represent a very high confidence that the query sequence belongs to the same protein family as the sequences use to create the domain model
non-specific hits
meet or exceed the RPS-BLAST threshold for statistical significance (default E-value cutoff of 0.01, or an E-value selected by user via the
advanced search options)
the domain superfamily to which the specific and non-specific hits belong
multi-domain models that were computationally detected and are likely to contain multiple single domains
Retrieve proteins that contain one or more of the domains present in the query sequence, using the Conserved Domain Architecture Retrieval Tool
(CDART).
Modify your query to search against a different database and/or use advanced search options