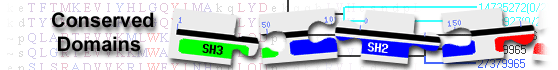


kinesin-like protein KIF1A isoform 1 [Homo sapiens]
Protein Classification

List of domain hits
| Name | Accession | Description | Interval | E-value | ||||||
| KISc_KIF1A_KIF1B | cd01365 | Kinesin motor domain, KIF1_like proteins; Kinesin motor domain, KIF1_like proteins. KIF1A ... |
4-361 | 0e+00 | ||||||
Kinesin motor domain, KIF1_like proteins; Kinesin motor domain, KIF1_like proteins. KIF1A (Unc104) transports synaptic vesicles to the nerve terminal, KIF1B has been implicated in transport of mitochondria. Both proteins are expressed in neurons. This catalytic (head) domain has ATPase activity and belongs to the larger group of P-loop NTPases. Kinesins are microtubule-dependent molecular motors that play important roles in intracellular transport and in cell division. In most kinesins, the motor domain is found at the N-terminus (N-type). N-type kinesins are (+) end-directed motors, i.e. they transport cargo towards the (+) end of the microtubule. In contrast to the majority of dimeric kinesins, most KIF1A/Unc104 kinesins are monomeric motors. A lysine-rich loop in KIF1A binds to the negatively charged C-terminus of tubulin and compensates for the lack of a second motor domain, allowing KIF1A to move processively. : Pssm-ID: 276816 [Multi-domain] Cd Length: 361 Bit Score: 679.46 E-value: 0e+00
|
||||||||||
| Kinesin_assoc | pfam16183 | Kinesin-associated; |
358-524 | 3.92e-95 | ||||||
Kinesin-associated; : Pssm-ID: 465047 [Multi-domain] Cd Length: 177 Bit Score: 304.46 E-value: 3.92e-95
|
||||||||||
| FHA_KIF1A | cd22726 | forkhead associated (FHA) domain found in kinesin-like protein KIF1A; KIF1A, also called ... |
500-614 | 8.81e-82 | ||||||
forkhead associated (FHA) domain found in kinesin-like protein KIF1A; KIF1A, also called axonal transporter of synaptic vesicles (ATSV), microtubule-based motor KIF1A, Unc-104- and KIF1A-related protein, or Unc-104, is an axonal transporter of synaptic vesicles, which is mutated in hereditary sensory and autonomic neuropathy type 2. It is also required for neuronal dense core vesicle (DCV) transport to dendritic spines and axons. The calcium-dependent interaction with CALM1 increases vesicle motility, and interaction with the scaffolding proteins PPFIA2 and TANC2 recruits DCVs to synaptic sites. The FHA domain is a small phosphopeptide recognition module, but this group may lack the conserved residues that are required for binding phosphothreonine. : Pssm-ID: 438778 [Multi-domain] Cd Length: 115 Bit Score: 263.71 E-value: 8.81e-82
|
||||||||||
| DUF3694 | pfam12473 | Kinesin protein; This domain family is found in eukaryotes, and is typically between 131 and ... |
1250-1397 | 2.37e-59 | ||||||
Kinesin protein; This domain family is found in eukaryotes, and is typically between 131 and 151 amino acids in length. The family is found in association with pfam00225, pfam00498. There is a single completely conserved residue W that may be functionally important. : Pssm-ID: 463599 Cd Length: 149 Bit Score: 200.89 E-value: 2.37e-59
|
||||||||||
| PH_KIFIA_KIFIB | cd01233 | KIFIA and KIFIB protein pleckstrin homology (PH) domain; The kinesin-3 family motors KIFIA ... |
1672-1774 | 1.50e-54 | ||||||
KIFIA and KIFIB protein pleckstrin homology (PH) domain; The kinesin-3 family motors KIFIA (Caenorhabditis elegans homolog unc-104) and KIFIB transport synaptic vesicle precursors that contain synaptic vesicle proteins, such as synaptophysin, synaptotagmin and the small GTPase RAB3A, but they do not transport organelles that contain plasma membrane proteins. They have a N-terminal motor domain, followed by a coiled-coil domain, and a C-terminal PH domain. KIF1A adopts a monomeric form in vitro, but acts as a processive dimer in vivo. KIF1B has alternatively spliced isoforms distinguished by the presence or absence of insertion sequences in the conserved amino-terminal region of the protein; this results in their different motor activities. KIF1A and KIF1B bind to RAB3 proteins through the adaptor protein mitogen-activated protein kinase (MAPK) -activating death domain (MADD; also calledDENN), which was first identified as a RAB3 guanine nucleotide exchange factor (GEF). PH domains have diverse functions, but in general are involved in targeting proteins to the appropriate cellular location or in the interaction with a binding partner. They share little sequence conservation, but all have a common fold, which is electrostatically polarized. Less than 10% of PH domains bind phosphoinositide phosphates (PIPs) with high affinity and specificity. PH domains are distinguished from other PIP-binding domains by their specific high-affinity binding to PIPs with two vicinal phosphate groups: PtdIns(3,4)P2, PtdIns(4,5)P2 or PtdIns(3,4,5)P3 which results in targeting some PH domain proteins to the plasma membrane. A few display strong specificity in lipid binding. Any specificity is usually determined by loop regions or insertions in the N-terminus of the domain, which are not conserved across all PH domains. PH domains are found in cellular signaling proteins such as serine/threonine kinase, tyrosine kinases, regulators of G-proteins, endocytotic GTPases, adaptors, as well as cytoskeletal associated molecules and in lipid associated enzymes. : Pssm-ID: 269939 Cd Length: 103 Bit Score: 185.10 E-value: 1.50e-54
|
||||||||||
| KIF1B | pfam12423 | Kinesin protein 1B; This domain family is found in eukaryotes, and is approximately 50 amino ... |
814-861 | 8.17e-15 | ||||||
Kinesin protein 1B; This domain family is found in eukaryotes, and is approximately 50 amino acids in length. The family is found in association with pfam00225, pfam00498. KIF1B is an anterograde motor for transport of mitochondria in axons of neuronal cells. : Pssm-ID: 463574 Cd Length: 43 Bit Score: 69.94 E-value: 8.17e-15
|
||||||||||
| ERM_helical super family | cl48646 | Ezrin/radixin/moesin, alpha-helical domain; The ERM family consists of three closely-related ... |
633-686 | 7.55e-03 | ||||||
Ezrin/radixin/moesin, alpha-helical domain; The ERM family consists of three closely-related proteins, ezrin, radixin and moesin. Ezrin was first identified as a constituent of microvilli, radixin as a barbed, end-capping actin-modulating protein from isolated junctional fractions, and moesin as a heparin binding protein. A tumour suppressor molecule responsible for neurofibromatosis type 2 (NF2) is highly similar to ERM proteins and has been designated merlin (moesin-ezrin-radixin-like protein). ERM molecules contain 3 domains, an N-terminal globular domain, an extended alpha-helical domain and a charged C-terminal domain (pfam00769). Ezrin, radixin and merlin also contain a polyproline linker region between the helical and C-terminal domains. The N-terminal domain is highly conserved and is also found in merlin, band 4.1 proteins and members of the band 4.1 superfamily, designated the FERM domain. ERM proteins crosslink actin filaments with plasma membranes. They co-localize with CD44 at actin filament plasma membrane interaction sites, associating with CD44 via their N-terminal domains and with actin filaments via their C-terminal domains. This is the alpha-helical domain, which is involved in intramolecular masking of protein-protein interaction sites, regulating the activity of this proteins. The actual alignment was detected with superfamily member pfam20492: Pssm-ID: 466641 [Multi-domain] Cd Length: 120 Bit Score: 38.36 E-value: 7.55e-03
|
||||||||||
| Name | Accession | Description | Interval | E-value | ||||||
| KISc_KIF1A_KIF1B | cd01365 | Kinesin motor domain, KIF1_like proteins; Kinesin motor domain, KIF1_like proteins. KIF1A ... |
4-361 | 0e+00 | ||||||
Kinesin motor domain, KIF1_like proteins; Kinesin motor domain, KIF1_like proteins. KIF1A (Unc104) transports synaptic vesicles to the nerve terminal, KIF1B has been implicated in transport of mitochondria. Both proteins are expressed in neurons. This catalytic (head) domain has ATPase activity and belongs to the larger group of P-loop NTPases. Kinesins are microtubule-dependent molecular motors that play important roles in intracellular transport and in cell division. In most kinesins, the motor domain is found at the N-terminus (N-type). N-type kinesins are (+) end-directed motors, i.e. they transport cargo towards the (+) end of the microtubule. In contrast to the majority of dimeric kinesins, most KIF1A/Unc104 kinesins are monomeric motors. A lysine-rich loop in KIF1A binds to the negatively charged C-terminus of tubulin and compensates for the lack of a second motor domain, allowing KIF1A to move processively. Pssm-ID: 276816 [Multi-domain] Cd Length: 361 Bit Score: 679.46 E-value: 0e+00
|
||||||||||
| KISc | smart00129 | Kinesin motor, catalytic domain. ATPase; Microtubule-dependent molecular motors that play ... |
5-361 | 1.62e-152 | ||||||
Kinesin motor, catalytic domain. ATPase; Microtubule-dependent molecular motors that play important roles in intracellular transport of organelles and in cell division. Pssm-ID: 214526 [Multi-domain] Cd Length: 335 Bit Score: 471.29 E-value: 1.62e-152
|
||||||||||
| Kinesin | pfam00225 | Kinesin motor domain; |
11-354 | 8.62e-152 | ||||||
Kinesin motor domain; Pssm-ID: 459720 [Multi-domain] Cd Length: 326 Bit Score: 468.98 E-value: 8.62e-152
|
||||||||||
| Kinesin_assoc | pfam16183 | Kinesin-associated; |
358-524 | 3.92e-95 | ||||||
Kinesin-associated; Pssm-ID: 465047 [Multi-domain] Cd Length: 177 Bit Score: 304.46 E-value: 3.92e-95
|
||||||||||
| FHA_KIF1A | cd22726 | forkhead associated (FHA) domain found in kinesin-like protein KIF1A; KIF1A, also called ... |
500-614 | 8.81e-82 | ||||||
forkhead associated (FHA) domain found in kinesin-like protein KIF1A; KIF1A, also called axonal transporter of synaptic vesicles (ATSV), microtubule-based motor KIF1A, Unc-104- and KIF1A-related protein, or Unc-104, is an axonal transporter of synaptic vesicles, which is mutated in hereditary sensory and autonomic neuropathy type 2. It is also required for neuronal dense core vesicle (DCV) transport to dendritic spines and axons. The calcium-dependent interaction with CALM1 increases vesicle motility, and interaction with the scaffolding proteins PPFIA2 and TANC2 recruits DCVs to synaptic sites. The FHA domain is a small phosphopeptide recognition module, but this group may lack the conserved residues that are required for binding phosphothreonine. Pssm-ID: 438778 [Multi-domain] Cd Length: 115 Bit Score: 263.71 E-value: 8.81e-82
|
||||||||||
| KIP1 | COG5059 | Kinesin-like protein [Cytoskeleton]; |
5-366 | 4.15e-81 | ||||||
Kinesin-like protein [Cytoskeleton]; Pssm-ID: 227392 [Multi-domain] Cd Length: 568 Bit Score: 279.32 E-value: 4.15e-81
|
||||||||||
| PLN03188 | PLN03188 | kinesin-12 family protein; Provisional |
1-381 | 2.12e-74 | ||||||
kinesin-12 family protein; Provisional Pssm-ID: 215621 [Multi-domain] Cd Length: 1320 Bit Score: 273.73 E-value: 2.12e-74
|
||||||||||
| DUF3694 | pfam12473 | Kinesin protein; This domain family is found in eukaryotes, and is typically between 131 and ... |
1250-1397 | 2.37e-59 | ||||||
Kinesin protein; This domain family is found in eukaryotes, and is typically between 131 and 151 amino acids in length. The family is found in association with pfam00225, pfam00498. There is a single completely conserved residue W that may be functionally important. Pssm-ID: 463599 Cd Length: 149 Bit Score: 200.89 E-value: 2.37e-59
|
||||||||||
| PH_KIFIA_KIFIB | cd01233 | KIFIA and KIFIB protein pleckstrin homology (PH) domain; The kinesin-3 family motors KIFIA ... |
1672-1774 | 1.50e-54 | ||||||
KIFIA and KIFIB protein pleckstrin homology (PH) domain; The kinesin-3 family motors KIFIA (Caenorhabditis elegans homolog unc-104) and KIFIB transport synaptic vesicle precursors that contain synaptic vesicle proteins, such as synaptophysin, synaptotagmin and the small GTPase RAB3A, but they do not transport organelles that contain plasma membrane proteins. They have a N-terminal motor domain, followed by a coiled-coil domain, and a C-terminal PH domain. KIF1A adopts a monomeric form in vitro, but acts as a processive dimer in vivo. KIF1B has alternatively spliced isoforms distinguished by the presence or absence of insertion sequences in the conserved amino-terminal region of the protein; this results in their different motor activities. KIF1A and KIF1B bind to RAB3 proteins through the adaptor protein mitogen-activated protein kinase (MAPK) -activating death domain (MADD; also calledDENN), which was first identified as a RAB3 guanine nucleotide exchange factor (GEF). PH domains have diverse functions, but in general are involved in targeting proteins to the appropriate cellular location or in the interaction with a binding partner. They share little sequence conservation, but all have a common fold, which is electrostatically polarized. Less than 10% of PH domains bind phosphoinositide phosphates (PIPs) with high affinity and specificity. PH domains are distinguished from other PIP-binding domains by their specific high-affinity binding to PIPs with two vicinal phosphate groups: PtdIns(3,4)P2, PtdIns(4,5)P2 or PtdIns(3,4,5)P3 which results in targeting some PH domain proteins to the plasma membrane. A few display strong specificity in lipid binding. Any specificity is usually determined by loop regions or insertions in the N-terminus of the domain, which are not conserved across all PH domains. PH domains are found in cellular signaling proteins such as serine/threonine kinase, tyrosine kinases, regulators of G-proteins, endocytotic GTPases, adaptors, as well as cytoskeletal associated molecules and in lipid associated enzymes. Pssm-ID: 269939 Cd Length: 103 Bit Score: 185.10 E-value: 1.50e-54
|
||||||||||
| KIF1B | pfam12423 | Kinesin protein 1B; This domain family is found in eukaryotes, and is approximately 50 amino ... |
814-861 | 8.17e-15 | ||||||
Kinesin protein 1B; This domain family is found in eukaryotes, and is approximately 50 amino acids in length. The family is found in association with pfam00225, pfam00498. KIF1B is an anterograde motor for transport of mitochondria in axons of neuronal cells. Pssm-ID: 463574 Cd Length: 43 Bit Score: 69.94 E-value: 8.17e-15
|
||||||||||
| PH | smart00233 | Pleckstrin homology domain; Domain commonly found in eukaryotic signalling proteins. The ... |
1677-1770 | 2.79e-11 | ||||||
Pleckstrin homology domain; Domain commonly found in eukaryotic signalling proteins. The domain family possesses multiple functions including the abilities to bind inositol phosphates, and various proteins. PH domains have been found to possess inserted domains (such as in PLC gamma, syntrophins) and to be inserted within other domains. Mutations in Brutons tyrosine kinase (Btk) within its PH domain cause X-linked agammaglobulinaemia (XLA) in patients. Point mutations cluster into the positively charged end of the molecule around the predicted binding site for phosphatidylinositol lipids. Pssm-ID: 214574 [Multi-domain] Cd Length: 102 Bit Score: 61.80 E-value: 2.79e-11
|
||||||||||
| PH | pfam00169 | PH domain; PH stands for pleckstrin homology. |
1677-1770 | 3.77e-10 | ||||||
PH domain; PH stands for pleckstrin homology. Pssm-ID: 459697 [Multi-domain] Cd Length: 105 Bit Score: 58.73 E-value: 3.77e-10
|
||||||||||
| FHA | COG1716 | Forkhead associated (FHA) domain, binds pSer, pThr, pTyr [Signal transduction mechanisms]; |
520-603 | 1.26e-07 | ||||||
Forkhead associated (FHA) domain, binds pSer, pThr, pTyr [Signal transduction mechanisms]; Pssm-ID: 441322 [Multi-domain] Cd Length: 96 Bit Score: 51.11 E-value: 1.26e-07
|
||||||||||
| FHA | pfam00498 | FHA domain; The FHA (Forkhead-associated) domain is a phosphopeptide binding motif. |
525-593 | 3.25e-07 | ||||||
FHA domain; The FHA (Forkhead-associated) domain is a phosphopeptide binding motif. Pssm-ID: 459831 [Multi-domain] Cd Length: 66 Bit Score: 49.11 E-value: 3.25e-07
|
||||||||||
| FHA | smart00240 | Forkhead associated domain; Found in eukaryotic and prokaryotic proteins. Putative nuclear ... |
525-581 | 4.74e-06 | ||||||
Forkhead associated domain; Found in eukaryotic and prokaryotic proteins. Putative nuclear signalling domain. Pssm-ID: 214578 [Multi-domain] Cd Length: 52 Bit Score: 45.25 E-value: 4.74e-06
|
||||||||||
| ERM_helical | pfam20492 | Ezrin/radixin/moesin, alpha-helical domain; The ERM family consists of three closely-related ... |
633-686 | 7.55e-03 | ||||||
Ezrin/radixin/moesin, alpha-helical domain; The ERM family consists of three closely-related proteins, ezrin, radixin and moesin. Ezrin was first identified as a constituent of microvilli, radixin as a barbed, end-capping actin-modulating protein from isolated junctional fractions, and moesin as a heparin binding protein. A tumour suppressor molecule responsible for neurofibromatosis type 2 (NF2) is highly similar to ERM proteins and has been designated merlin (moesin-ezrin-radixin-like protein). ERM molecules contain 3 domains, an N-terminal globular domain, an extended alpha-helical domain and a charged C-terminal domain (pfam00769). Ezrin, radixin and merlin also contain a polyproline linker region between the helical and C-terminal domains. The N-terminal domain is highly conserved and is also found in merlin, band 4.1 proteins and members of the band 4.1 superfamily, designated the FERM domain. ERM proteins crosslink actin filaments with plasma membranes. They co-localize with CD44 at actin filament plasma membrane interaction sites, associating with CD44 via their N-terminal domains and with actin filaments via their C-terminal domains. This is the alpha-helical domain, which is involved in intramolecular masking of protein-protein interaction sites, regulating the activity of this proteins. Pssm-ID: 466641 [Multi-domain] Cd Length: 120 Bit Score: 38.36 E-value: 7.55e-03
|
||||||||||
| Name | Accession | Description | Interval | E-value | ||||||
| KISc_KIF1A_KIF1B | cd01365 | Kinesin motor domain, KIF1_like proteins; Kinesin motor domain, KIF1_like proteins. KIF1A ... |
4-361 | 0e+00 | ||||||
Kinesin motor domain, KIF1_like proteins; Kinesin motor domain, KIF1_like proteins. KIF1A (Unc104) transports synaptic vesicles to the nerve terminal, KIF1B has been implicated in transport of mitochondria. Both proteins are expressed in neurons. This catalytic (head) domain has ATPase activity and belongs to the larger group of P-loop NTPases. Kinesins are microtubule-dependent molecular motors that play important roles in intracellular transport and in cell division. In most kinesins, the motor domain is found at the N-terminus (N-type). N-type kinesins are (+) end-directed motors, i.e. they transport cargo towards the (+) end of the microtubule. In contrast to the majority of dimeric kinesins, most KIF1A/Unc104 kinesins are monomeric motors. A lysine-rich loop in KIF1A binds to the negatively charged C-terminus of tubulin and compensates for the lack of a second motor domain, allowing KIF1A to move processively. Pssm-ID: 276816 [Multi-domain] Cd Length: 361 Bit Score: 679.46 E-value: 0e+00
|
||||||||||
| KISc | cd00106 | Kinesin motor domain; Kinesin motor domain. This catalytic (head) domain has ATPase activity ... |
5-352 | 3.75e-154 | ||||||
Kinesin motor domain; Kinesin motor domain. This catalytic (head) domain has ATPase activity and belongs to the larger group of P-loop NTPases. Kinesins are microtubule-dependent molecular motors that play important roles in intracellular transport and in cell division. In most kinesins, the motor domain is found at the N-terminus (N-type), in some its is found in the middle (M-type), or C-terminal (C-type). N-type and M-type kinesins are (+) end-directed motors, while C-type kinesins are (-) end-directed motors, i.e. they transport cargo towards the (-) end of the microtubule. Kinesin motor domains hydrolyze ATP at a rate of about 80 per second, and move along the microtubule at a speed of about 6400 Angstroms per second. To achieve that, kinesin head groups work in pairs. Upon replacing ADP with ATP, a kinesin motor domain increases its affinity for microtubule binding and locks in place. Also, the neck linker binds to the motor domain, which repositions the other head domain through the coiled-coil domain close to a second tubulin dimer, about 80 Angstroms along the microtubule. Meanwhile, ATP hydrolysis takes place, and when the second head domain binds to the microtubule, the first domain again replaces ADP with ATP, triggering a conformational change that pulls the first domain forward. Pssm-ID: 276812 [Multi-domain] Cd Length: 326 Bit Score: 475.21 E-value: 3.75e-154
|
||||||||||
| KISc | smart00129 | Kinesin motor, catalytic domain. ATPase; Microtubule-dependent molecular motors that play ... |
5-361 | 1.62e-152 | ||||||
Kinesin motor, catalytic domain. ATPase; Microtubule-dependent molecular motors that play important roles in intracellular transport of organelles and in cell division. Pssm-ID: 214526 [Multi-domain] Cd Length: 335 Bit Score: 471.29 E-value: 1.62e-152
|
||||||||||
| Kinesin | pfam00225 | Kinesin motor domain; |
11-354 | 8.62e-152 | ||||||
Kinesin motor domain; Pssm-ID: 459720 [Multi-domain] Cd Length: 326 Bit Score: 468.98 E-value: 8.62e-152
|
||||||||||
| KISc_KIF3 | cd01371 | Kinesin motor domain, kinesins II or KIF3_like proteins; Kinesin motor domain, kinesins II or ... |
5-354 | 1.03e-113 | ||||||
Kinesin motor domain, kinesins II or KIF3_like proteins; Kinesin motor domain, kinesins II or KIF3_like proteins. Subgroup of kinesins, which form heterotrimers composed of 2 kinesins and one non-motor accessory subunit. Kinesins II play important roles in ciliary transport, and have been implicated in neuronal transport, melanosome transport, the secretory pathway, and mitosis. This catalytic (head) domain has ATPase activity and belongs to the larger group of P-loop NTPases. Kinesins are microtubule-dependent molecular motors that play important roles in intracellular transport and in cell division. In this group the motor domain is found at the N-terminus (N-type). N-type kinesins are (+) end-directed motors, i.e. they transport cargo towards the (+) end of the microtubule. Kinesin motor domains hydrolyze ATP at a rate of about 80 per second, and move along the microtubule at a speed of about 6400 Angstroms per second. To achieve that, kinesin head groups work in pairs. Upon replacing ADP with ATP, a kinesin motor domain increases its affinity for microtubule binding and locks in place. Also, the neck linker binds to the motor domain, which repositions the other head domain through the coiled-coil domain close to a second tubulin dimer, about 80 Angstroms along the microtubule. Meanwhile, ATP hydrolysis takes place, and when the second head domain binds to the microtubule, the first domain again replaces ADP with ATP, triggering a conformational change that pulls the first domain forward. Pssm-ID: 276822 [Multi-domain] Cd Length: 334 Bit Score: 364.09 E-value: 1.03e-113
|
||||||||||
| KISc_KHC_KIF5 | cd01369 | Kinesin motor domain, kinesin heavy chain (KHC) or KIF5-like subgroup; Kinesin motor domain, ... |
5-354 | 2.92e-109 | ||||||
Kinesin motor domain, kinesin heavy chain (KHC) or KIF5-like subgroup; Kinesin motor domain, kinesin heavy chain (KHC) or KIF5-like subgroup. Members of this group have been associated with organelle transport. This catalytic (head) domain has ATPase activity and belongs to the larger group of P-loop NTPases. Kinesins are microtubule-dependent molecular motors that play important roles in intracellular transport and in cell division. In most kinesins, the motor domain is found at the N-terminus (N-type). N-type kinesins are (+) end-directed motors, i.e. they transport cargo towards the (+) end of the microtubule. Kinesin motor domains hydrolyze ATP at a rate of about 80 per second, and move along the microtubule at a speed of about 6400 Angstroms per second. To achieve that, kinesin head groups work in pairs. Upon replacing ADP with ATP, a kinesin motor domain increases its affinity for microtubule binding and locks in place. Also, the neck linker binds to the motor domain, which repositions the other head domain through the coiled-coil domain close to a second tubulin dimer, about 80 Angstroms along the microtubule. Meanwhile, ATP hydrolysis takes place, and when the second head domain binds to the microtubule, the first domain again replaces ADP with ATP, triggering a conformational change that pulls the first domain forward. Pssm-ID: 276820 [Multi-domain] Cd Length: 325 Bit Score: 351.25 E-value: 2.92e-109
|
||||||||||
| KISc_KIF4 | cd01372 | Kinesin motor domain, KIF4-like subfamily; Kinesin motor domain, KIF4-like subfamily. Members ... |
5-355 | 3.75e-104 | ||||||
Kinesin motor domain, KIF4-like subfamily; Kinesin motor domain, KIF4-like subfamily. Members of this group seem to perform a variety of functions, and have been implicated in neuronal organelle transport and chromosome segregation during mitosis. This catalytic (head) domain has ATPase activity and belongs to the larger group of P-loop NTPases. Kinesins are microtubule-dependent molecular motors that play important roles in intracellular transport and in cell division. In most kinesins, the motor domain is found at the N-terminus (N-type). N-type kinesins are (+) end-directed motors, i.e. they transport cargo towards the (+) end of the microtubule. Kinesin motor domains hydrolyze ATP at a rate of about 80 per second, and move along the microtubule at a speed of about 6400 Angstroms per second. To achieve that, kinesin head groups work in pairs. Upon replacing ADP with ATP, a kinesin motor domain increases its affinity for microtubule binding and locks in place. Also, the neck linker binds to the motor domain, which repositions the other head domain through the coiled-coil domain close to a second tubulin dimer, about 80 Angstroms along the microtubule. Meanwhile, ATP hydrolysis takes place, and when the second head domain binds to the microtubule, the first domain again replaces ADP with ATP, triggering a conformational change that pulls the first domain forward. Pssm-ID: 276823 [Multi-domain] Cd Length: 341 Bit Score: 337.38 E-value: 3.75e-104
|
||||||||||
| KISc_CENP_E | cd01374 | Kinesin motor domain, CENP-E/KIP2-like subgroup; Kinesin motor domain, CENP-E/KIP2-like ... |
5-354 | 3.52e-103 | ||||||
Kinesin motor domain, CENP-E/KIP2-like subgroup; Kinesin motor domain, CENP-E/KIP2-like subgroup, involved in chromosome movement and/or spindle elongation during mitosis. This catalytic (head) domain has ATPase activity and belongs to the larger group of P-loop NTPases. Kinesins are microtubule-dependent molecular motors that play important roles in intracellular transport and in cell division. In most kinesins, the motor domain is found at the N-terminus (N-type). N-type kinesins are (+) end-directed motors, i.e. they transport cargo towards the (+) end of the microtubule. Kinesin motor domains hydrolyze ATP at a rate of about 80 per second, and move along the microtubule at a speed of about 6400 Angstroms per second. To achieve that, kinesin head groups work in pairs. Upon replacing ADP with ATP, a kinesin motor domain increases its affinity for microtubule binding and locks in place. Also, the neck linker binds to the motor domain, which repositions the other head domain through the coiled-coil domain close to a second tubulin dimer, about 80 Angstroms along the microtubule. Meanwhile, ATP hydrolysis takes place, and when the second head domain binds to the microtubule, the first domain again replaces ADP with ATP, triggering a conformational change that pulls the first domain forward. Pssm-ID: 276825 [Multi-domain] Cd Length: 321 Bit Score: 333.53 E-value: 3.52e-103
|
||||||||||
| KISc_KIP3_like | cd01370 | Kinesin motor domain, KIP3-like subgroup; Kinesin motor domain, KIP3-like subgroup. The yeast ... |
5-354 | 5.13e-102 | ||||||
Kinesin motor domain, KIP3-like subgroup; Kinesin motor domain, KIP3-like subgroup. The yeast kinesin KIP3 plays a role in positioning the mitotic spindle. This catalytic (head) domain has ATPase activity and belongs to the larger group of P-loop NTPases. Kinesins are microtubule-dependent molecular motors that play important roles in intracellular transport and in cell division. In most kinesins, the motor domain is found at the N-terminus (N-type). N-type kinesins are (+) end-directed motors, i.e. they transport cargo towards the (+) end of the microtubule. Kinesin motor domains hydrolyze ATP at a rate of about 80 per second, and move along the microtubule at a speed of about 6400 Angstroms per second. To achieve that, kinesin head groups work in pairs. Upon replacing ADP with ATP, a kinesin motor domain increases its affinity for microtubule binding and locks in place. Also, the neck linker binds to the motor domain, which repositions the other head domain through the coiled-coil domain close to a second tubulin dimer, about 80 Angstroms along the microtubule. Meanwhile, ATP hydrolysis takes place, and when the second head domain binds to the microtubule, the first domain again replaces ADP with ATP, triggering a conformational change that pulls the first domain forward. Pssm-ID: 276821 [Multi-domain] Cd Length: 345 Bit Score: 331.23 E-value: 5.13e-102
|
||||||||||
| KISc_C_terminal | cd01366 | Kinesin motor domain, KIFC2/KIFC3/ncd-like carboxy-terminal kinesins; Kinesin motor domain, ... |
11-356 | 5.16e-100 | ||||||
Kinesin motor domain, KIFC2/KIFC3/ncd-like carboxy-terminal kinesins; Kinesin motor domain, KIFC2/KIFC3/ncd-like carboxy-terminal kinesins. Ncd is a spindle motor protein necessary for chromosome segregation in meiosis. KIFC2/KIFC3-like kinesins have been implicated in motility of the Golgi apparatus as well as dentritic and axonal transport in neurons. This catalytic (head) domain has ATPase activity and belongs to the larger group of P-loop NTPases. Kinesins are microtubule-dependent molecular motors that play important roles in intracellular transport and in cell division. In this subgroup the motor domain is found at the C-terminus (C-type). C-type kinesins are (-) end-directed motors, i.e. they transport cargo towards the (-) end of the microtubule. Kinesin motor domains hydrolyze ATP at a rate of about 80 per second, and move along the microtubule at a speed of about 6400 Angstroms per second. To achieve that, kinesin head groups work in pairs. Upon replacing ADP with ATP, a kinesin motor domain increases its affinity for microtubule binding and locks in place. Also, the neck linker binds to the motor domain, which repositions the other head domain through the coiled-coil domain close to a second tubulin dimer, about 80 Angstroms along the microtubule. Meanwhile, ATP hydrolysis takes place, and when the second head domain binds to the microtubule, the first domain again replaces ADP with ATP, triggering a conformational change that pulls the first domain forward. Pssm-ID: 276817 [Multi-domain] Cd Length: 329 Bit Score: 324.93 E-value: 5.16e-100
|
||||||||||
| KISc_KLP2_like | cd01373 | Kinesin motor domain, KIF15-like subgroup; Kinesin motor domain, KIF15-like subgroup. Members ... |
6-363 | 1.18e-95 | ||||||
Kinesin motor domain, KIF15-like subgroup; Kinesin motor domain, KIF15-like subgroup. Members of this subgroup seem to play a role in mitosis and meiosis. This catalytic (head) domain has ATPase activity and belongs to the larger group of P-loop NTPases. Kinesins are microtubule-dependent molecular motors that play important roles in intracellular transport and in cell division. In most kinesins, the motor domain is found at the N-terminus (N-type). N-type kinesins are (+) end-directed motors, i.e. they transport cargo towards the (+) end of the microtubule. Kinesin motor domains hydrolyze ATP at a rate of about 80 per second, and move along the microtubule at a speed of about 6400 Angstroms per second. To achieve that, kinesin head groups work in pairs. Upon replacing ADP with ATP, a kinesin motor domain increases its affinity for microtubule binding and locks in place. Also, the neck linker binds to the motor domain, which repositions the other head domain through the coiled-coil domain close to a second tubulin dimer, about 80 Angstroms along the microtubule. Meanwhile, ATP hydrolysis takes place, and when the second head domain binds to the microtubule, the first domain again replaces ADP with ATP, triggering a conformational change that pulls the first domain forward. Pssm-ID: 276824 [Multi-domain] Cd Length: 347 Bit Score: 313.29 E-value: 1.18e-95
|
||||||||||
| Kinesin_assoc | pfam16183 | Kinesin-associated; |
358-524 | 3.92e-95 | ||||||
Kinesin-associated; Pssm-ID: 465047 [Multi-domain] Cd Length: 177 Bit Score: 304.46 E-value: 3.92e-95
|
||||||||||
| KISc_BimC_Eg5 | cd01364 | Kinesin motor domain, BimC/Eg5 spindle pole proteins; Kinesin motor domain, BimC/Eg5 spindle ... |
3-363 | 3.16e-93 | ||||||
Kinesin motor domain, BimC/Eg5 spindle pole proteins; Kinesin motor domain, BimC/Eg5 spindle pole proteins, participate in spindle assembly and chromosome segregation during cell division. This catalytic (head) domain has ATPase activity and belongs to the larger group of P-loop NTPases. Kinesins are microtubule-dependent molecular motors that play important roles in intracellular transport and in cell division. In most kinesins, the motor domain is found at the N-terminus (N-type), N-type kinesins are (+) end-directed motors, i.e. they transport cargo towards the (+) end of the microtubule. Kinesin motor domains hydrolyze ATP at a rate of about 80 per second, and move along the microtubule at a speed of about 6400 Angstroms per second. To achieve that, kinesin head groups work in pairs. Upon replacing ADP with ATP, a kinesin motor domain increases its affinity for microtubule binding and locks in place. Also, the neck linker binds to the motor domain, which repositions the other head domain through the coiled-coil domain close to a second tubulin dimer, about 80 Angstroms along the microtubule. Meanwhile, ATP hydrolysis takes place, and when the second head domain binds to the microtubule, the first domain again replaces ADP with ATP, triggering a conformational change that pulls the first domain forward. Pssm-ID: 276815 [Multi-domain] Cd Length: 353 Bit Score: 306.56 E-value: 3.16e-93
|
||||||||||
| FHA_KIF1A | cd22726 | forkhead associated (FHA) domain found in kinesin-like protein KIF1A; KIF1A, also called ... |
500-614 | 8.81e-82 | ||||||
forkhead associated (FHA) domain found in kinesin-like protein KIF1A; KIF1A, also called axonal transporter of synaptic vesicles (ATSV), microtubule-based motor KIF1A, Unc-104- and KIF1A-related protein, or Unc-104, is an axonal transporter of synaptic vesicles, which is mutated in hereditary sensory and autonomic neuropathy type 2. It is also required for neuronal dense core vesicle (DCV) transport to dendritic spines and axons. The calcium-dependent interaction with CALM1 increases vesicle motility, and interaction with the scaffolding proteins PPFIA2 and TANC2 recruits DCVs to synaptic sites. The FHA domain is a small phosphopeptide recognition module, but this group may lack the conserved residues that are required for binding phosphothreonine. Pssm-ID: 438778 [Multi-domain] Cd Length: 115 Bit Score: 263.71 E-value: 8.81e-82
|
||||||||||
| KIP1 | COG5059 | Kinesin-like protein [Cytoskeleton]; |
5-366 | 4.15e-81 | ||||||
Kinesin-like protein [Cytoskeleton]; Pssm-ID: 227392 [Multi-domain] Cd Length: 568 Bit Score: 279.32 E-value: 4.15e-81
|
||||||||||
| PLN03188 | PLN03188 | kinesin-12 family protein; Provisional |
1-381 | 2.12e-74 | ||||||
kinesin-12 family protein; Provisional Pssm-ID: 215621 [Multi-domain] Cd Length: 1320 Bit Score: 273.73 E-value: 2.12e-74
|
||||||||||
| KISc_KIF2_like | cd01367 | Kinesin motor domain, KIF2-like group; Kinesin motor domain, KIF2-like group. KIF2 is a ... |
6-352 | 4.39e-74 | ||||||
Kinesin motor domain, KIF2-like group; Kinesin motor domain, KIF2-like group. KIF2 is a protein expressed in neurons, which has been associated with axonal transport and neuron development; alternative splice forms have been implicated in lysosomal translocation. This catalytic (head) domain has ATPase activity and belongs to the larger group of P-loop NTPases. Kinesins are microtubule-dependent molecular motors that play important roles in intracellular transport and in cell division. In this subgroup the motor domain is found in the middle (M-type) of the protein chain. M-type kinesins are (+) end-directed motors, i.e. they transport cargo towards the (+) end of the microtubule. Kinesin motor domains hydrolyze ATP at a rate of about 80 per second, and move along the microtubule at a speed of about 6400 Angstroms per second (KIF2 may be slower). To achieve that, kinesin head groups work in pairs. Upon replacing ADP with ATP, a kinesin motor domain increases its affinity for microtubule binding and locks in place. Also, the neck linker binds to the motor domain, which repositions the other head domain through the coiled-coil domain close to a second tubulin dimer, about 80 Angstroms along the microtubule. Meanwhile, ATP hydrolysis takes place, and when the second head domain binds to the microtubule, the first domain again replaces ADP with ATP, triggering a conformational change that pulls the first domain forward. Pssm-ID: 276818 [Multi-domain] Cd Length: 328 Bit Score: 250.29 E-value: 4.39e-74
|
||||||||||
| KISc_KIF23_like | cd01368 | Kinesin motor domain, KIF23-like subgroup; Kinesin motor domain, KIF23-like subgroup. Members ... |
6-352 | 6.69e-72 | ||||||
Kinesin motor domain, KIF23-like subgroup; Kinesin motor domain, KIF23-like subgroup. Members of this group may play a role in mitosis. This catalytic (head) domain has ATPase activity and belongs to the larger group of P-loop NTPases. Kinesins are microtubule-dependent molecular motors that play important roles in intracellular transport and in cell division. In most kinesins, the motor domain is found at the N-terminus (N-type). N-type kinesins are (+) end-directed motors, i.e. they transport cargo towards the (+) end of the microtubule. Kinesin motor domains hydrolyze ATP at a rate of about 80 per second, and move along the microtubule at a speed of about 6400 Angstroms per second. To achieve that, kinesin head groups work in pairs. Upon replacing ADP with ATP, a kinesin motor domain increases its affinity for microtubule binding and locks in place. Also, the neck linker binds to the motor domain, which repositions the other head domain through the coiled-coil domain close to a second tubulin dimer, about 80 Angstroms along the microtubule. Meanwhile, ATP hydrolysis takes place, and when the second head domain binds to the microtubule, the first domain again replaces ADP with ATP, triggering a conformational change that pulls the first domain forward. Pssm-ID: 276819 [Multi-domain] Cd Length: 345 Bit Score: 244.61 E-value: 6.69e-72
|
||||||||||
| KISc_KID_like | cd01376 | Kinesin motor domain, KIF22/Kid-like subgroup; Kinesin motor domain, KIF22/Kid-like subgroup. ... |
6-352 | 2.57e-70 | ||||||
Kinesin motor domain, KIF22/Kid-like subgroup; Kinesin motor domain, KIF22/Kid-like subgroup. Members of this group might play a role in regulating chromosomal movement along microtubules in mitosis. This catalytic (head) domain has ATPase activity and belongs to the larger group of P-loop NTPases. Kinesins are microtubule-dependent molecular motors that play important roles in intracellular transport and in cell division. In most kinesins, the motor domain is found at the N-terminus (N-type). N-type kinesins are (+) end-directed motors, i.e. they transport cargo towards the (+) end of the microtubule. Kinesin motor domains hydrolyze ATP at a rate of about 80 per second, and move along the microtubule at a speed of about 6400 Angstroms per second. To achieve that, kinesin head groups work in pairs. Upon replacing ADP with ATP, a kinesin motor domain increases its affinity for microtubule binding and locks in place. Also, the neck linker binds to the motor domain, which repositions the other head domain through the coiled-coil domain close to a second tubulin dimer, about 80 Angstroms along the microtubule. Meanwhile, ATP hydrolysis takes place, and when the second head domain binds to the microtubule, the first domain again replaces ADP with ATP, triggering a conformational change that pulls the first domain forward. Pssm-ID: 276827 [Multi-domain] Cd Length: 319 Bit Score: 239.33 E-value: 2.57e-70
|
||||||||||
| FHA_KIF1B | cd22727 | forkhead associated (FHA) domain found in kinesin-like protein KIF1B; KIF1B, also called Klp, ... |
499-608 | 1.08e-66 | ||||||
forkhead associated (FHA) domain found in kinesin-like protein KIF1B; KIF1B, also called Klp, is a motor for anterograde transport of mitochondria. It has a microtubule plus end-directed motility. Isoform 1 mediates the transport of synaptic vesicles in neuronal cells, while isoform 2 is required for induction of neuronal apoptosis. The FHA domain is a small phosphopeptide recognition module, but this group may lack the conserved residues that are required for binding phosphothreonine. Pssm-ID: 438779 [Multi-domain] Cd Length: 110 Bit Score: 220.29 E-value: 1.08e-66
|
||||||||||
| KISc_KIF9_like | cd01375 | Kinesin motor domain, KIF9-like subgroup; Kinesin motor domain, KIF9-like subgroup; might play ... |
5-352 | 9.63e-64 | ||||||
Kinesin motor domain, KIF9-like subgroup; Kinesin motor domain, KIF9-like subgroup; might play a role in cell shape remodeling. This catalytic (head) domain has ATPase activity and belongs to the larger group of P-loop NTPases. Kinesins are microtubule-dependent molecular motors that play important roles in intracellular transport and in cell division. In most kinesins, the motor domain is found at the N-terminus (N-type). N-type kinesins are (+) end-directed motors, i.e. they transport cargo towards the (+) end of the microtubule. Kinesin motor domains hydrolyze ATP at a rate of about 80 per second, and move along the microtubule at a speed of about 6400 Angstroms per second. To achieve that, kinesin head groups work in pairs. Upon replacing ADP with ATP, a kinesin motor domain increases its affinity for microtubule binding and locks in place. Also, the neck linker binds to the motor domain, which repositions the other head domain through the coiled-coil domain close to a second tubulin dimer, about 80 Angstroms along the microtubule. Meanwhile, ATP hydrolysis takes place, and when the second head domain binds to the microtubule, the first domain again replaces ADP with ATP, triggering a conformational change that pulls the first domain forward. Pssm-ID: 276826 [Multi-domain] Cd Length: 334 Bit Score: 220.91 E-value: 9.63e-64
|
||||||||||
| FHA_KIF1 | cd22705 | forkhead associated (FHA) domain found in the kinesin-like protein KIF1 family; The KIF1 ... |
500-605 | 3.11e-63 | ||||||
forkhead associated (FHA) domain found in the kinesin-like protein KIF1 family; The KIF1 family includes KIF1A, KIF1B, and KIF1C. KIF1A, also called axonal transporter of synaptic vesicles (ATSV), microtubule-based motor KIF1A, Unc-104- and KIF1A-related protein, or Unc-104, is an axonal transporter of synaptic vesicles, which is mutated in hereditary sensory and autonomic neuropathy type 2. It is also required for neuronal dense core vesicle (DCV) transport to dendritic spines and axons. The calcium-dependent interaction with CALM1 increases vesicle motility, and interaction with the scaffolding proteins PPFIA2 and TANC2 recruits DCVs to synaptic sites. KIF1B, also called Klp, is a motor for anterograde transport of mitochondria. It has a microtubule plus end-directed motility. Isoform 1 mediates the transport of synaptic vesicles in neuronal cells, while isoform 2 is required for induction of neuronal apoptosis. KIF1C is a new kinesin-like protein involved in vesicle transport from the Golgi apparatus to the endoplasmic reticulum. It has a microtubule plus end-directed motility. The FHA domain is a small phosphopeptide recognition module, but this group may lack the conserved residues that are required for binding phosphothreonine. Pssm-ID: 438757 [Multi-domain] Cd Length: 101 Bit Score: 210.17 E-value: 3.11e-63
|
||||||||||
| DUF3694 | pfam12473 | Kinesin protein; This domain family is found in eukaryotes, and is typically between 131 and ... |
1250-1397 | 2.37e-59 | ||||||
Kinesin protein; This domain family is found in eukaryotes, and is typically between 131 and 151 amino acids in length. The family is found in association with pfam00225, pfam00498. There is a single completely conserved residue W that may be functionally important. Pssm-ID: 463599 Cd Length: 149 Bit Score: 200.89 E-value: 2.37e-59
|
||||||||||
| FHA_KIF1C | cd22728 | forkhead associated (FHA) domain found in kinesin-like protein KIF1C; KIF1C is a new ... |
500-605 | 7.79e-57 | ||||||
forkhead associated (FHA) domain found in kinesin-like protein KIF1C; KIF1C is a new kinesin-like protein involved in vesicle transport from the Golgi apparatus to the endoplasmic reticulum. It has a microtubule plus end-directed motility. The FHA domain is a small phosphopeptide recognition module, but this group may lack the conserved residues that are required for binding phosphothreonine. Pssm-ID: 438780 [Multi-domain] Cd Length: 102 Bit Score: 192.01 E-value: 7.79e-57
|
||||||||||
| PH_KIFIA_KIFIB | cd01233 | KIFIA and KIFIB protein pleckstrin homology (PH) domain; The kinesin-3 family motors KIFIA ... |
1672-1774 | 1.50e-54 | ||||||
KIFIA and KIFIB protein pleckstrin homology (PH) domain; The kinesin-3 family motors KIFIA (Caenorhabditis elegans homolog unc-104) and KIFIB transport synaptic vesicle precursors that contain synaptic vesicle proteins, such as synaptophysin, synaptotagmin and the small GTPase RAB3A, but they do not transport organelles that contain plasma membrane proteins. They have a N-terminal motor domain, followed by a coiled-coil domain, and a C-terminal PH domain. KIF1A adopts a monomeric form in vitro, but acts as a processive dimer in vivo. KIF1B has alternatively spliced isoforms distinguished by the presence or absence of insertion sequences in the conserved amino-terminal region of the protein; this results in their different motor activities. KIF1A and KIF1B bind to RAB3 proteins through the adaptor protein mitogen-activated protein kinase (MAPK) -activating death domain (MADD; also calledDENN), which was first identified as a RAB3 guanine nucleotide exchange factor (GEF). PH domains have diverse functions, but in general are involved in targeting proteins to the appropriate cellular location or in the interaction with a binding partner. They share little sequence conservation, but all have a common fold, which is electrostatically polarized. Less than 10% of PH domains bind phosphoinositide phosphates (PIPs) with high affinity and specificity. PH domains are distinguished from other PIP-binding domains by their specific high-affinity binding to PIPs with two vicinal phosphate groups: PtdIns(3,4)P2, PtdIns(4,5)P2 or PtdIns(3,4,5)P3 which results in targeting some PH domain proteins to the plasma membrane. A few display strong specificity in lipid binding. Any specificity is usually determined by loop regions or insertions in the N-terminus of the domain, which are not conserved across all PH domains. PH domains are found in cellular signaling proteins such as serine/threonine kinase, tyrosine kinases, regulators of G-proteins, endocytotic GTPases, adaptors, as well as cytoskeletal associated molecules and in lipid associated enzymes. Pssm-ID: 269939 Cd Length: 103 Bit Score: 185.10 E-value: 1.50e-54
|
||||||||||
| Motor_domain | cd01363 | Myosin and Kinesin motor domain; Myosin and Kinesin motor domain. These ATPases belong to the ... |
8-284 | 2.13e-45 | ||||||
Myosin and Kinesin motor domain; Myosin and Kinesin motor domain. These ATPases belong to the P-loop NTPase family and provide the driving force in myosin and kinesin mediated processes. Some of the names do not match with what is given in the sequence list. This is because they are based on the current nomenclature by Kollmar/Sebe-Pedros. Pssm-ID: 276814 [Multi-domain] Cd Length: 170 Bit Score: 161.74 E-value: 2.13e-45
|
||||||||||
| FHA_KIF14 | cd22707 | forkhead associated (FHA) domain found in kinesin-like protein KIF14 and similar proteins; ... |
499-606 | 1.30e-36 | ||||||
forkhead associated (FHA) domain found in kinesin-like protein KIF14 and similar proteins; KIF14 is a microtubule motor protein that binds to microtubules with high affinity through each tubulin heterodimer and has an ATPase activity. It plays a role in many processes like cell division, cytokinesis and in cell proliferation and apoptosis. KIF14 is a potential oncogene and is involved in the metastasis of various cancers. Mutations of KIF14 cause primary microcephaly by impairing cytokinesis. The FHA domain is a small phosphopeptide recognition module, but this group may lack the conserved residues that are required for binding phosphothreonine. Pssm-ID: 438759 [Multi-domain] Cd Length: 108 Bit Score: 134.32 E-value: 1.30e-36
|
||||||||||
| FHA_KIF16 | cd22708 | forkhead associated (FHA) domain found in the kinesin-like protein KIF16 family; The KIF16 ... |
499-606 | 1.92e-36 | ||||||
forkhead associated (FHA) domain found in the kinesin-like protein KIF16 family; The KIF16 family includes StARD9/KIF16A and KIF16B. StARD9, also called START domain-containing protein 9, or kinesin-like protein KIF16A, is a microtubule-dependent motor protein required for spindle pole assembly during mitosis. It is required to stabilize the pericentriolar material (PCM). KIF16B, also called sorting nexin-23, is a plus end-directed microtubule-dependent motor protein involved in endosome transport and receptor recycling and degradation. It regulates the plus end motility of early endosomes and the balance between recycling and degradation of receptors such as EGF receptor (EGFR) and FGF receptor (FGFR). It regulates the Golgi to endosome transport of FGFR-containing vesicles during early development, a key process for developing basement membrane and epiblast and primitive endoderm lineages during early postimplantation development. The FHA domain is a small phosphopeptide recognition module, but this group may lack the conserved residues that are required for binding phosphothreonine. Pssm-ID: 438760 [Multi-domain] Cd Length: 109 Bit Score: 133.94 E-value: 1.92e-36
|
||||||||||
| FHA_KIF28P | cd22709 | forkhead associated (FHA) domain found in kinesin-like protein KIF28P and similar proteins; ... |
501-606 | 2.12e-35 | ||||||
forkhead associated (FHA) domain found in kinesin-like protein KIF28P and similar proteins; KIF28P, also called kinesin-like protein 6 (KLP6), is a microtubule-dependent motor protein required for mitochondrion morphology and transport of mitochondria in neuronal cells. The FHA domain is a small phosphopeptide recognition module, but this group may lack the conserved residues that are required for binding phosphothreonine. Pssm-ID: 438761 [Multi-domain] Cd Length: 102 Bit Score: 130.41 E-value: 2.12e-35
|
||||||||||
| FHA_KIF16A_STARD9 | cd22731 | forkhead associated (FHA) domain found in StAR-related lipid transfer protein 9 (StARD9); ... |
493-615 | 1.69e-32 | ||||||
forkhead associated (FHA) domain found in StAR-related lipid transfer protein 9 (StARD9); StARD9, also called START domain-containing protein 9, or kinesin-like protein KIF16A, is a microtubule-dependent motor protein required for spindle pole assembly during mitosis. It is required to stabilize the pericentriolar material (PCM). The FHA domain is a small phosphopeptide recognition module, but this group may lack the conserved residues that are required for binding phosphothreonine. Pssm-ID: 438783 [Multi-domain] Cd Length: 119 Bit Score: 122.96 E-value: 1.69e-32
|
||||||||||
| FHA_KIF13 | cd22706 | forkhead associated (FHA) domain found in the kinesin-like protein KIF13 family; The KIF13 ... |
503-606 | 2.38e-31 | ||||||
forkhead associated (FHA) domain found in the kinesin-like protein KIF13 family; The KIF13 family includes KIF13A and KIF13B. KIF13A, also called kinesin-like protein RBKIN, is a plus end-directed microtubule-dependent motor protein involved in intracellular transport and in regulating various processes such as mannose-6-phosphate receptor (M6PR) transport to the plasma membrane, endosomal sorting during melanosome biogenesis, and cytokinesis. It mediates the transport of M6PR-containing vesicles from trans-Golgi network to the plasma membrane via direct interaction with the AP-1 complex. During melanosome maturation, KIF13A is required for delivering melanogenic enzymes from recycling endosomes to nascent melanosomes by creating peripheral recycling endosomal subdomains in melanocytes. It is also required for the abscission step in cytokinesis: it mediates translocation of ZFYVE26, and possibly TTC19, to the midbody during cytokinesis. KIF13B, also called kinesin-like protein GAKIN, is a novel kinesin-like protein that associates with the human homolog of the Drosophila discs large tumor suppressor in T lymphocytes. It is involved in reorganization of the cortical cytoskeleton. It regulates axon formation by promoting the formation of extra axons. KIF13B may be functionally important for the intracellular trafficking of membrane-associated guanylate kinase homologs (MAGUKs) and associated protein complexes. The FHA domain is a small phosphopeptide recognition module, but this group may lack the conserved residues that are required for binding phosphothreonine. Pssm-ID: 438758 [Multi-domain] Cd Length: 101 Bit Score: 118.94 E-value: 2.38e-31
|
||||||||||
| FHA_KIF16B | cd22732 | forkhead associated (FHA) domain found in kinesin-like protein KIF16B; KIF16B, also called ... |
493-614 | 2.05e-27 | ||||||
forkhead associated (FHA) domain found in kinesin-like protein KIF16B; KIF16B, also called sorting nexin-23, is a plus end-directed microtubule-dependent motor protein involved in endosome transport and receptor recycling and degradation. It regulates the plus end motility of early endosomes and the balance between recycling and degradation of receptors such as EGF receptor (EGFR) and FGF receptor (FGFR). It regulates the Golgi to endosome transport of FGFR-containing vesicles during early development, a key process for developing basement membrane and epiblast and primitive endoderm lineages during early postimplantation development. The FHA domain is a small phosphopeptide recognition module, but this group may lack the conserved residues that are required for binding phosphothreonine. Pssm-ID: 438784 [Multi-domain] Cd Length: 117 Bit Score: 108.48 E-value: 2.05e-27
|
||||||||||
| FHA_KIF13A | cd22729 | forkhead associated (FHA) domain found in kinesin-like protein KIF13A; KIF13A, also called ... |
503-613 | 3.50e-20 | ||||||
forkhead associated (FHA) domain found in kinesin-like protein KIF13A; KIF13A, also called kinesin-like protein RBKIN, is a plus end-directed microtubule-dependent motor protein involved in intracellular transport and in regulating various processes such as mannose-6-phosphate receptor (M6PR) transport to the plasma membrane, endosomal sorting during melanosome biogenesis, and cytokinesis. It mediates the transport of M6PR-containing vesicles from trans-Golgi network to the plasma membrane via direct interaction with the AP-1 complex. During melanosome maturation, KIF13A is required for delivering melanogenic enzymes from recycling endosomes to nascent melanosomes by creating peripheral recycling endosomal subdomains in melanocytes. It is also required for the abscission step in cytokinesis: it mediates translocation of ZFYVE26, and possibly TTC19, to the midbody during cytokinesis. The FHA domain is a small phosphopeptide recognition module, but this group may lack the conserved residues that are required for binding phosphothreonine. Pssm-ID: 438781 [Multi-domain] Cd Length: 109 Bit Score: 87.25 E-value: 3.50e-20
|
||||||||||
| FHA_KIF13B | cd22730 | forkhead associated (FHA) domain found in kinesin-like protein KIF13B; KIF13B, also called ... |
503-606 | 2.57e-19 | ||||||
forkhead associated (FHA) domain found in kinesin-like protein KIF13B; KIF13B, also called kinesin-like protein GAKIN, is a novel kinesin-like protein that associates with the human homolog of the Drosophila discs large tumor suppressor in T lymphocytes. It is involved in reorganization of the cortical cytoskeleton. It regulates axon formation by promoting the formation of extra axons. KIF13B may be functionally important for the intracellular trafficking of membrane-associated guanylate kinase homologs (MAGUKs) and associated protein complexes. The FHA domain is a small phosphopeptide recognition module, but this group may lack the conserved residues that are required for binding phosphothreonine. Pssm-ID: 438782 [Multi-domain] Cd Length: 99 Bit Score: 84.58 E-value: 2.57e-19
|
||||||||||
| FHA_AFDN | cd22711 | forkhead associated (FHA) domain found in afadin and similar proteins; Afadin, also called ... |
501-606 | 4.20e-17 | ||||||
forkhead associated (FHA) domain found in afadin and similar proteins; Afadin, also called ALL1-fused gene from chromosome 6 protein, protein AF-6, Afadin adherens junction formation factor, or MLLT4, is a nectin- and actin-filament-binding protein that connects nectin to the actin cytoskeleton. It is essential for the organization of adherens junctions. It may play a key role in the organization of epithelial structures of the embryonic ectoderm. The FHA domain is a small phosphopeptide recognition module, but this group may lack the conserved residues that are required for binding phosphothreonine. Pssm-ID: 438763 [Multi-domain] Cd Length: 106 Bit Score: 78.52 E-value: 4.20e-17
|
||||||||||
| FHA_PHLB1 | cd22713 | forkhead associated (FHA) domain found in pleckstrin homology-like domain family B member 1 ... |
487-611 | 1.85e-16 | ||||||
forkhead associated (FHA) domain found in pleckstrin homology-like domain family B member 1 (PHLDB1) and similar proteins; PHLDB1, also called protein LL5-alpha (LL5A), acts as an insulin-responsive protein that enhances Akt activation. PHLDB1 contains a pleckstrin homology domain, which binds phosphatidylinositol PI(3,4)P(2), PI(3,5)P(2), and PI(3,4,5)P(3), as well as a Forkhead-associated (FHA) domain and coiled coil regions. The FHA domain is a small phosphopeptide recognition module, but this group may lack the conserved residues that are required for binding phosphothreonine. Pssm-ID: 438765 Cd Length: 120 Bit Score: 76.98 E-value: 1.85e-16
|
||||||||||
| KIF1B | pfam12423 | Kinesin protein 1B; This domain family is found in eukaryotes, and is approximately 50 amino ... |
814-861 | 8.17e-15 | ||||||
Kinesin protein 1B; This domain family is found in eukaryotes, and is approximately 50 amino acids in length. The family is found in association with pfam00225, pfam00498. KIF1B is an anterograde motor for transport of mitochondria in axons of neuronal cells. Pssm-ID: 463574 Cd Length: 43 Bit Score: 69.94 E-value: 8.17e-15
|
||||||||||
| Microtub_bd | pfam16796 | Microtubule binding; This motor homology domain binds microtubules and lacks an ATP-binding ... |
5-158 | 9.41e-15 | ||||||
Microtubule binding; This motor homology domain binds microtubules and lacks an ATP-binding site. Pssm-ID: 465274 [Multi-domain] Cd Length: 144 Bit Score: 73.02 E-value: 9.41e-15
|
||||||||||
| PH | smart00233 | Pleckstrin homology domain; Domain commonly found in eukaryotic signalling proteins. The ... |
1677-1770 | 2.79e-11 | ||||||
Pleckstrin homology domain; Domain commonly found in eukaryotic signalling proteins. The domain family possesses multiple functions including the abilities to bind inositol phosphates, and various proteins. PH domains have been found to possess inserted domains (such as in PLC gamma, syntrophins) and to be inserted within other domains. Mutations in Brutons tyrosine kinase (Btk) within its PH domain cause X-linked agammaglobulinaemia (XLA) in patients. Point mutations cluster into the positively charged end of the molecule around the predicted binding site for phosphatidylinositol lipids. Pssm-ID: 214574 [Multi-domain] Cd Length: 102 Bit Score: 61.80 E-value: 2.79e-11
|
||||||||||
| FHA | cd00060 | forkhead associated (FHA) domain superfamily; Forkhead-associated (FHA) domains are small ... |
502-603 | 9.21e-11 | ||||||
forkhead associated (FHA) domain superfamily; Forkhead-associated (FHA) domains are small phosphopeptide recognition modules mostly found in eubacteria and eukaryotes. It is about 95-120 residues long that fold into an 11-stranded beta-sandwich. FHA domains can mediate the recognition of phosphorylated and non-phosphorylated substrates, as well as protein oligomerization. They specifically recognize threonine phosphorylation (pThr) accompanying activation of protein serine/threonine kinases. FHA domains show diverse ligand specificity. They may recognize the pTXXD motif, the pTXXI/L motif, and TQ clusters (singly and multiply phosphorylated). In eukaryotes, FHA superfamily members include forkhead-type transcription factors, as well as other signaling proteins, such as many regulatory proteins, kinases, phosphatases, motor proteins called kinesins, and metabolic enzymes. Many of them localize to the nucleus, where they participate in establishing or maintaining cell cycle checkpoints, DNA repair, or transcriptional regulation. FHA domains play important roles in human diseases, particularly in relation to DNA damage responses and cancers. In bacteria, FHA domain-containing proteins may participate in injection of viral proteins into host cells, transmembrane transporters, and cell division. FHA domain-containing proteins rarely include more than one copy of the domain. The only exception in eukaryotes is the checkpoint kinase Rad53 from Saccharomyces cerevisiae, which harbors two FHA domains (FHA1 and FHA2) flanking a central kinase domain. The two FHA domains recognize different phosphorylated targets and function independently from one another. In contrast, Mycobacterium tuberculosis ABC transporter Rv1747 contains two FHA domains but only one of them is essential for protein function. Pssm-ID: 438714 [Multi-domain] Cd Length: 92 Bit Score: 59.98 E-value: 9.21e-11
|
||||||||||
| PH | pfam00169 | PH domain; PH stands for pleckstrin homology. |
1677-1770 | 3.77e-10 | ||||||
PH domain; PH stands for pleckstrin homology. Pssm-ID: 459697 [Multi-domain] Cd Length: 105 Bit Score: 58.73 E-value: 3.77e-10
|
||||||||||
| FHA_RADIL | cd22733 | forkhead associated (FHA) domain found in Ras-associating and dilute domain-containing protein ... |
499-606 | 1.19e-08 | ||||||
forkhead associated (FHA) domain found in Ras-associating and dilute domain-containing protein (Radil); Radil acts as an important small GTPase Rap1 effector required for cell spreading and migration. It regulates neutrophil adhesion and motility through linking Rap1 to beta2-integrin activation. It contains an FHA domain. The FHA domain is a small phosphopeptide recognition module, but this group may lack the conserved residues that are required for binding phosphothreonine. Pssm-ID: 438785 Cd Length: 113 Bit Score: 54.80 E-value: 1.19e-08
|
||||||||||
| PH1_PH_fungal | cd13298 | Fungal proteins Pleckstrin homology (PH) domain, repeat 1; The functions of these fungal ... |
1677-1770 | 1.56e-08 | ||||||
Fungal proteins Pleckstrin homology (PH) domain, repeat 1; The functions of these fungal proteins are unknown, but they all contain 2 PH domains. This cd represents the first PH repeat. PH domains have diverse functions, but in general are involved in targeting proteins to the appropriate cellular location or in the interaction with a binding partner. They share little sequence conservation, but all have a common fold, which is electrostatically polarized. Less than 10% of PH domains bind phosphoinositide phosphates (PIPs) with high affinity and specificity. PH domains are distinguished from other PIP-binding domains by their specific high-affinity binding to PIPs with two vicinal phosphate groups: PtdIns(3,4)P2, PtdIns(4,5)P2 or PtdIns(3,4,5)P3 which results in targeting some PH domain proteins to the plasma membrane. A few display strong specificity in lipid binding. Any specificity is usually determined by loop regions or insertions in the N-terminus of the domain, which are not conserved across all PH domains. PH domains are found in cellular signaling proteins such as serine/threonine kinase, tyrosine kinases, regulators of G-proteins, endocytotic GTPases, adaptors, as well as cytoskeletal associated molecules and in lipid associated enzymes. Pssm-ID: 270110 Cd Length: 106 Bit Score: 54.17 E-value: 1.56e-08
|
||||||||||
| FHA | COG1716 | Forkhead associated (FHA) domain, binds pSer, pThr, pTyr [Signal transduction mechanisms]; |
520-603 | 1.26e-07 | ||||||
Forkhead associated (FHA) domain, binds pSer, pThr, pTyr [Signal transduction mechanisms]; Pssm-ID: 441322 [Multi-domain] Cd Length: 96 Bit Score: 51.11 E-value: 1.26e-07
|
||||||||||
| PH_TBC1D2A | cd01265 | TBC1 domain family member 2A pleckstrin homology (PH) domain; TBC1D2A (also called PARIS-1 ... |
1681-1766 | 1.91e-07 | ||||||
TBC1 domain family member 2A pleckstrin homology (PH) domain; TBC1D2A (also called PARIS-1/Prostate antigen recognized and identified by SEREX 1 and ARMUS) contains a PH domain and a TBC-type GTPase catalytic domain. TBC1D2A integrates signaling between Arf6, Rac1, and Rab7 during junction disassembly. Activated Rac1 recruits TBC1D2A to locally inactivate Rab7 via its C-terminal TBC/RabGAP domain and facilitate E-cadherin degradation in lysosomes. The TBC1D2A PH domain mediates localization at cell-cell contacts and coprecipitates with cadherin complexes. PH domains have diverse functions, but in general are involved in targeting proteins to the appropriate cellular location or in the interaction with a binding partner. They share little sequence conservation, but all have a common fold, which is electrostatically polarized. Less than 10% of PH domains bind phosphoinositide phosphates (PIPs) with high affinity and specificity. PH domains are distinguished from other PIP-binding domains by their specific high-affinity binding to PIPs with two vicinal phosphate groups: PtdIns(3,4)P2, PtdIns(4,5)P2 or PtdIns(3,4,5)P3 which results in targeting some PH domain proteins to the plasma membrane. A few display strong specificity in lipid binding. Any specificity is usually determined by loop regions or insertions in the N-terminus of the domain, which are not conserved across all PH domains. PH domains are found in cellular signaling proteins such as serine/threonine kinase, tyrosine kinases, regulators of G-proteins, endocytotic GTPases, adaptors, as well as cytoskeletal associated molecules and in lipid associated enzymes. Pssm-ID: 269966 Cd Length: 102 Bit Score: 50.78 E-value: 1.91e-07
|
||||||||||
| FHA_RADIL-like | cd22712 | forkhead associated (FHA) domain found in the Ras-associating and dilute domain-containing ... |
500-606 | 2.05e-07 | ||||||
forkhead associated (FHA) domain found in the Ras-associating and dilute domain-containing protein (Radil)-like family; The Radil-like family includes Radil and Ras-interacting protein 1 (Rain). Radil acts as an important small GTPase Rap1 effector required for cell spreading and migration. It regulates neutrophil adhesion and motility by linking Rap1 to beta2-integrin activation. Rain, also called Rasip1, is an endothelial-specific Ras-interacting protein required for the proper formation of vascular structures that develop via both vasculogenesis and angiogenesis. It acts as a critical and vascular-specific regulator of GTPase signaling, cell architecture, and adhesion, which is essential for endothelial cell morphogenesis and blood vessel tubulogenesis. Rain interacts with Ras in a GTP-dependent manner and may serve as an effector for endomembrane-associated Ras. Both Radil and Rain contain an FHA domain. The FHA domain is a small phosphopeptide recognition module, but this group may lack the conserved residues that are required for binding phosphothreonine. Pssm-ID: 438764 [Multi-domain] Cd Length: 120 Bit Score: 51.15 E-value: 2.05e-07
|
||||||||||
| FHA | pfam00498 | FHA domain; The FHA (Forkhead-associated) domain is a phosphopeptide binding motif. |
525-593 | 3.25e-07 | ||||||
FHA domain; The FHA (Forkhead-associated) domain is a phosphopeptide binding motif. Pssm-ID: 459831 [Multi-domain] Cd Length: 66 Bit Score: 49.11 E-value: 3.25e-07
|
||||||||||
| PH | cd00821 | Pleckstrin homology (PH) domain; PH domains have diverse functions, but in general are ... |
1679-1769 | 3.79e-06 | ||||||
Pleckstrin homology (PH) domain; PH domains have diverse functions, but in general are involved in targeting proteins to the appropriate cellular location or in the interaction with a binding partner. They share little sequence conservation, but all have a common fold, which is electrostatically polarized. Less than 10% of PH domains bind phosphoinositide phosphates (PIPs) with high affinity and specificity. PH domains are distinguished from other PIP-binding domains by their specific high-affinity binding to PIPs with two vicinal phosphate groups: PtdIns(3,4)P2, PtdIns(4,5)P2 or PtdIns(3,4,5)P3 which results in targeting some PH domain proteins to the plasma membrane. A few display strong specificity in lipid binding. Any specificity is usually determined by loop regions or insertions in the N-terminus of the domain, which are not conserved across all PH domains. PH domains are found in cellular signaling proteins such as serine/threonine kinase, tyrosine kinases, regulators of G-proteins, endocytotic GTPases, adaptors, as well as cytoskeletal associated molecules and in lipid associated enzymes. Pssm-ID: 275388 [Multi-domain] Cd Length: 92 Bit Score: 46.77 E-value: 3.79e-06
|
||||||||||
| FHA | smart00240 | Forkhead associated domain; Found in eukaryotic and prokaryotic proteins. Putative nuclear ... |
525-581 | 4.74e-06 | ||||||
Forkhead associated domain; Found in eukaryotic and prokaryotic proteins. Putative nuclear signalling domain. Pssm-ID: 214578 [Multi-domain] Cd Length: 52 Bit Score: 45.25 E-value: 4.74e-06
|
||||||||||
| PH_TAAP2-like | cd13255 | Tandem PH-domain-containing protein 2 Pleckstrin homology (PH) domain; The binding of TAPP2 ... |
1674-1770 | 7.32e-06 | ||||||
Tandem PH-domain-containing protein 2 Pleckstrin homology (PH) domain; The binding of TAPP2 (also called PLEKHA2) adaptors to PtdIns(3,4)P(2), but not PI(3,4, 5)P3, function as negative regulators of insulin and PI3K signalling pathways (i.e. TAPP/utrophin/syntrophin complex). TAPP2 contains two sequential PH domains in which the C-terminal PH domain specifically binds PtdIns(3,4)P2 with high affinity. The N-terminal PH domain does not interact with any phosphoinositide tested. They also contain a C-terminal PDZ-binding motif that interacts with several PDZ-binding proteins, including PTPN13 (known previously as PTPL1 or FAP-1) as well as the scaffolding proteins MUPP1 (multiple PDZ-domain-containing protein 1), syntrophin and utrophin. The members here are most sequence similar to TAPP2 proteins, but may not be actual TAPP2 proteins. PH domains have diverse functions, but in general are involved in targeting proteins to the appropriate cellular location or in the interaction with a binding partner. They share little sequence conservation, but all have a common fold, which is electrostatically polarized. Less than 10% of PH domains bind phosphoinositide phosphates (PIPs) with high affinity and specificity. PH domains are distinguished from other PIP-binding domains by their specific high-affinity binding to PIPs with two vicinal phosphate groups: PtdIns(3,4)P2, PtdIns(4,5)P2 or PtdIns(3,4,5)P3 which results in targeting some PH domain proteins to the plasma membrane. A few display strong specificity in lipid binding. Any specificity is usually determined by loop regions or insertions in the N-terminus of the domain, which are not conserved across all PH domains. PH domains are found in cellular signaling proteins such as serine/threonine kinase, tyrosine kinases, regulators of G-proteins, endocytotic GTPases, adaptors, as well as cytoskeletal associated molecules and in lipid associated enzymes. Pssm-ID: 270075 Cd Length: 110 Bit Score: 46.64 E-value: 7.32e-06
|
||||||||||
| PH_DAPP1 | cd10573 | Dual Adaptor for Phosphotyrosine and 3-Phosphoinositides Pleckstrin homology (PH) domain; ... |
1675-1765 | 3.85e-05 | ||||||
Dual Adaptor for Phosphotyrosine and 3-Phosphoinositides Pleckstrin homology (PH) domain; DAPP1 (also known as PHISH/3' phosphoinositide-interacting SH2 domain-containing protein or Bam32) plays a role in B-cell activation and has potential roles in T-cell and mast cell function. DAPP1 promotes B cell receptor (BCR) induced activation of Rho GTPases Rac1 and Cdc42, which feed into mitogen-activated protein kinases (MAPK) activation pathways and affect cytoskeletal rearrangement. DAPP1can also regulate BCR-induced activation of extracellular signal-regulated kinase (ERK), and c-jun NH2-terminal kinase (JNK). DAPP1 contains an N-terminal SH2 domain and a C-terminal pleckstrin homology (PH) domain with a single tyrosine phosphorylation site located centrally. DAPP1 binds strongly to both PtdIns(3,4,5)P3 and PtdIns(3,4)P2. The PH domain is essential for plasma membrane recruitment of PI3K upon cell activation. PH domains have diverse functions, but in general are involved in targeting proteins to the appropriate cellular location or in the interaction with a binding partner. They share little sequence conservation, but all have a common fold, which is electrostatically polarized. Less than 10% of PH domains bind phosphoinositide phosphates (PIPs) with high affinity and specificity. PH domains are distinguished from other PIP-binding domains by their specific high-affinity binding to PIPs with two vicinal phosphate groups: PtdIns(3,4)P2, PtdIns(4,5)P2 or PtdIns(3,4,5)P3 which results in targeting some PH domain proteins to the plasma membrane. A few display strong specificity in lipid binding. Any specificity is usually determined by loop regions or insertions in the N-terminus of the domain, which are not conserved across all PH domains. PH domains are found in cellular signaling proteins such as serine/threonine kinase, tyrosine kinases, regulators of G-proteins, endocytotic GTPases, adaptors, as well as cytoskeletal associated molecules and in lipid associated enzymes. Pssm-ID: 269977 [Multi-domain] Cd Length: 96 Bit Score: 44.24 E-value: 3.85e-05
|
||||||||||
| KIP1 | COG5059 | Kinesin-like protein [Cytoskeleton]; |
93-285 | 2.01e-04 | ||||||
Kinesin-like protein [Cytoskeleton]; Pssm-ID: 227392 [Multi-domain] Cd Length: 568 Bit Score: 46.27 E-value: 2.01e-04
|
||||||||||
| PH_ACAP | cd13250 | ArfGAP with coiled-coil, ankyrin repeat and PH domains Pleckstrin homology (PH) domain; ACAP ... |
1679-1768 | 1.45e-03 | ||||||
ArfGAP with coiled-coil, ankyrin repeat and PH domains Pleckstrin homology (PH) domain; ACAP (also called centaurin beta) functions both as a Rab35 effector and as an Arf6-GTPase-activating protein (GAP) by which it controls actin remodeling and membrane trafficking. ACAP contain an NH2-terminal bin/amphiphysin/Rvs (BAR) domain, a phospholipid-binding domain, a PH domain, a GAP domain, and four ankyrin repeats. The AZAPs constitute a family of Arf GAPs that are characterized by an NH2-terminal pleckstrin homology (PH) domain and a central Arf GAP domain followed by two or more ankyrin repeats. On the basis of sequence and domain organization, the AZAP family is further subdivided into four subfamilies: 1) the ACAPs contain an NH2-terminal bin/amphiphysin/Rvs (BAR) domain (a phospholipid-binding domain that is thought to sense membrane curvature), a single PH domain followed by the GAP domain, and four ankyrin repeats; 2) the ASAPs also contain an NH2-terminal BAR domain, the tandem PH domain/GAP domain, three ankyrin repeats, two proline-rich regions, and a COOH-terminal Src homology 3 domain; 3) the AGAPs contain an NH2-terminal GTPase-like domain (GLD), a split PH domain, and the GAP domain followed by four ankyrin repeats; and 4) the ARAPs contain both an Arf GAP domain and a Rho GAP domain, as well as an NH2-terminal sterile-a motif (SAM), a proline-rich region, a GTPase-binding domain, and five PH domains. PMID 18003747 and 19055940 Centaurin can bind to phosphatidlyinositol (3,4,5)P3. PH domains have diverse functions, but in general are involved in targeting proteins to the appropriate cellular location or in the interaction with a binding partner. They share little sequence conservation, but all have a common fold, which is electrostatically polarized. Less than 10% of PH domains bind phosphoinositide phosphates (PIPs) with high affinity and specificity. PH domains are distinguished from other PIP-binding domains by their specific high-affinity binding to PIPs with two vicinal phosphate groups: PtdIns(3,4)P2, PtdIns(4,5)P2 or PtdIns(3,4,5)P3 which results in targeting some PH domain proteins to the plasma membrane. A few display strong specificity in lipid binding. Any specificity is usually determined by loop regions or insertions in the N-terminus of the domain, which are not conserved across all PH domains. PH domains are found in cellular signaling proteins such as serine/threonine kinase, tyrosine kinases, regulators of G-proteins, endocytotic GTPases, adaptors, as well as cytoskeletal associated molecules and in lipid associated enzymes. Pssm-ID: 270070 Cd Length: 98 Bit Score: 39.51 E-value: 1.45e-03
|
||||||||||
| FHA_MEK1-like | cd22670 | forkhead associated (FHA) domain found in Saccharomyces cerevisiae meiosis-specific serine ... |
514-605 | 4.26e-03 | ||||||
forkhead associated (FHA) domain found in Saccharomyces cerevisiae meiosis-specific serine/threonine-protein kinase MEK1 and similar proteins; MEK1 (EC 2.7.11.1), also known as MRE4, is a meiosis-specific protein kinase required for chromosome synapsis and meiotic recombination. The recruitment and activation of MEK1 require the phosphorylation of the chromosome axis protein Hop1 at Thr318 (pT318), which is necessary for recognition by the MEK1 FHA domain. The FHA domain is a small phosphopeptide recognition module. Pssm-ID: 438722 [Multi-domain] Cd Length: 105 Bit Score: 38.36 E-value: 4.26e-03
|
||||||||||
| FHA_Ki67 | cd22673 | forkhead associated (FHA) domain found in proliferation marker protein Ki-67 and similar ... |
520-603 | 4.43e-03 | ||||||
forkhead associated (FHA) domain found in proliferation marker protein Ki-67 and similar proteins; Ki-67, also called antigen identified by monoclonal antibody Ki-67, antigen KI-67, or antigen Ki67, acts as a biological surfactant to disperse mitotic chromosomes. It is required to maintain individual mitotic chromosomes dispersed in the cytoplasm following nuclear envelope disassembly. Ki-67 binds DNA with a preference for supercoiled DNA and AT-rich DNA. It may also play a role in chromatin organization. Ki-67 contains an FHA domain at its N-terminus. The FHA domain is a small phosphopeptide recognition module. Pssm-ID: 438725 [Multi-domain] Cd Length: 95 Bit Score: 38.35 E-value: 4.43e-03
|
||||||||||
| PH_ARHGAP21-like | cd01253 | ARHGAP21 and related proteins pleckstrin homology (PH) domain; ARHGAP family genes encode Rho ... |
1679-1768 | 5.34e-03 | ||||||
ARHGAP21 and related proteins pleckstrin homology (PH) domain; ARHGAP family genes encode Rho/Rac/Cdc42-like GTPase activating proteins with a RhoGAP domain. These proteins functions as a GTPase-activating protein (GAP) for RHOA and CDC42. ARHGAP21 controls the Arp2/3 complex and F-actin dynamics at the Golgi complex by regulating the activity of the small GTPase Cdc42. It is recruited to the Golgi by to GTPase, ARF1, through its PH domain and its helical motif. It is also required for CTNNA1 recruitment to adherens junctions. ARHGAP21 and it related proteins all contains a PH domain and a RhoGAP domain. Some of the members have additional N-terminal domains including PDZ, SH3, and SPEC. The ARHGAP21 PH domain interacts with the GTPbound forms of both ARF1 and ARF6 ARF-binding domain/ArfBD. The members here include: ARHGAP15, ARHGAP21, and ARHGAP23. PH domains have diverse functions, but in general are involved in targeting proteins to the appropriate cellular location or in the interaction with a binding partner. They share little sequence conservation, but all have a common fold, which is electrostatically polarized. Less than 10% of PH domains bind phosphoinositide phosphates (PIPs) with high affinity and specificity. PH domains are distinguished from other PIP-binding domains by their specific high-affinity binding to PIPs with two vicinal phosphate groups: PtdIns(3,4)P2, PtdIns(4,5)P2 or PtdIns(3,4,5)P3 which results in targeting some PH domain proteins to the plasma membrane. A few display strong specificity in lipid binding. Any specificity is usually determined by loop regions or insertions in the N-terminus of the domain, which are not conserved across all PH domains. PH domains are found in cellular signaling proteins such as serine/threonine kinase, tyrosine kinases, regulators of G-proteins, endocytotic GTPases, adaptors, as well as cytoskeletal associated molecules and in lipid associated enzymes. Pssm-ID: 269955 Cd Length: 113 Bit Score: 38.51 E-value: 5.34e-03
|
||||||||||
| ERM_helical | pfam20492 | Ezrin/radixin/moesin, alpha-helical domain; The ERM family consists of three closely-related ... |
633-686 | 7.55e-03 | ||||||
Ezrin/radixin/moesin, alpha-helical domain; The ERM family consists of three closely-related proteins, ezrin, radixin and moesin. Ezrin was first identified as a constituent of microvilli, radixin as a barbed, end-capping actin-modulating protein from isolated junctional fractions, and moesin as a heparin binding protein. A tumour suppressor molecule responsible for neurofibromatosis type 2 (NF2) is highly similar to ERM proteins and has been designated merlin (moesin-ezrin-radixin-like protein). ERM molecules contain 3 domains, an N-terminal globular domain, an extended alpha-helical domain and a charged C-terminal domain (pfam00769). Ezrin, radixin and merlin also contain a polyproline linker region between the helical and C-terminal domains. The N-terminal domain is highly conserved and is also found in merlin, band 4.1 proteins and members of the band 4.1 superfamily, designated the FERM domain. ERM proteins crosslink actin filaments with plasma membranes. They co-localize with CD44 at actin filament plasma membrane interaction sites, associating with CD44 via their N-terminal domains and with actin filaments via their C-terminal domains. This is the alpha-helical domain, which is involved in intramolecular masking of protein-protein interaction sites, regulating the activity of this proteins. Pssm-ID: 466641 [Multi-domain] Cd Length: 120 Bit Score: 38.36 E-value: 7.55e-03
|
||||||||||
| Blast search parameters | ||||
|
