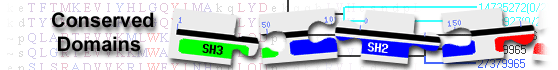DNA mismatch repair protein MutL; All proteins in this family for which the functions are ...
13-344
5.49e-143
DNA mismatch repair protein MutL; All proteins in this family for which the functions are known are involved in the process of generalized mismatch repair. This family is based on the phylogenomic analysis of JA Eisen (1999, Ph.D. Thesis, Stanford University). [DNA metabolism, DNA replication, recombination, and repair]
:
Pssm-ID: 273155 [Multi-domain] Cd Length: 312 Bit Score: 425.52 E-value: 5.49e-143
DNA mismatch repair protein MutL; All proteins in this family for which the functions are ...
13-344
5.49e-143
DNA mismatch repair protein MutL; All proteins in this family for which the functions are known are involved in the process of generalized mismatch repair. This family is based on the phylogenomic analysis of JA Eisen (1999, Ph.D. Thesis, Stanford University). [DNA metabolism, DNA replication, recombination, and repair]
Pssm-ID: 273155 [Multi-domain] Cd Length: 312 Bit Score: 425.52 E-value: 5.49e-143
Histidine kinase-like ATPase domain of DNA mismatch repair proteins Escherichia coli MutL, ...
22-205
1.78e-88
Histidine kinase-like ATPase domain of DNA mismatch repair proteins Escherichia coli MutL, human MutL homologs (MLH/ PMS), and related domains; This family includes the histidine kinase-like ATPase (HATPase) domains of Escherichia coli MutL, human MLH1 (mutL homolog 1), human PMS1 (PMS1 homolog 1, mismatch repair system component), human MLH3 (mutL homolog 3), and human PMS2 (PMS1 homolog 2, mismatch repair system component). MutL homologs (MLH/PMS) participate in MMR (DNA mismatch repair), and in addition have role(s) in DNA damage signaling and suppression of homologous recombination (recombination between partially homologous parental DNAs). The primary role of MutL in MMR is to mediate protein-protein interactions during mismatch recognition and strand removal; a ternary complex is formed between MutS, MutL, and the mismatched DNA, which activates the MutH endonuclease.
Pssm-ID: 340403 [Multi-domain] Cd Length: 188 Bit Score: 278.55 E-value: 1.78e-88
MutL C terminal dimerisation domain; MutL and MutS are key components of the DNA repair ...
678-832
9.92e-36
MutL C terminal dimerisation domain; MutL and MutS are key components of the DNA repair machinery that corrects replication errors. MutS recognises mispaired or unpaired bases in a DNA duplex and in the presence of ATP, recruits MutL to form a DNA signaling complex for repair. The N terminal region of MutL contains the ATPase domain and the C terminal is involved in dimerisation.
Pssm-ID: 214857 [Multi-domain] Cd Length: 140 Bit Score: 132.09 E-value: 9.92e-36
MutL C terminal dimerization domain; MutL and MutS are key components of the DNA repair ...
679-833
4.77e-28
MutL C terminal dimerization domain; MutL and MutS are key components of the DNA repair machinery that corrects replication errors. MutS recognizes mispaired or unpaired bases in a DNA duplex and in the presence of ATP, recruits MutL to form a DNA signaling complex for repair. The N terminal region of MutL contains the ATPase domain and the C terminal is involved in dimerization.
Pssm-ID: 430147 Cd Length: 145 Bit Score: 110.39 E-value: 4.77e-28
DNA mismatch repair protein, C-terminal domain; This family represents the C-terminal domain ...
229-359
8.25e-24
DNA mismatch repair protein, C-terminal domain; This family represents the C-terminal domain of the mutL/hexB/PMS1 family. This domain has a ribosomal S5 domain 2-like fold.
Pssm-ID: 426060 [Multi-domain] Cd Length: 117 Bit Score: 97.18 E-value: 8.25e-24
DNA mismatch repair protein MutL; All proteins in this family for which the functions are ...
13-344
5.49e-143
DNA mismatch repair protein MutL; All proteins in this family for which the functions are known are involved in the process of generalized mismatch repair. This family is based on the phylogenomic analysis of JA Eisen (1999, Ph.D. Thesis, Stanford University). [DNA metabolism, DNA replication, recombination, and repair]
Pssm-ID: 273155 [Multi-domain] Cd Length: 312 Bit Score: 425.52 E-value: 5.49e-143
Histidine kinase-like ATPase domain of DNA mismatch repair proteins Escherichia coli MutL, ...
22-205
1.78e-88
Histidine kinase-like ATPase domain of DNA mismatch repair proteins Escherichia coli MutL, human MutL homologs (MLH/ PMS), and related domains; This family includes the histidine kinase-like ATPase (HATPase) domains of Escherichia coli MutL, human MLH1 (mutL homolog 1), human PMS1 (PMS1 homolog 1, mismatch repair system component), human MLH3 (mutL homolog 3), and human PMS2 (PMS1 homolog 2, mismatch repair system component). MutL homologs (MLH/PMS) participate in MMR (DNA mismatch repair), and in addition have role(s) in DNA damage signaling and suppression of homologous recombination (recombination between partially homologous parental DNAs). The primary role of MutL in MMR is to mediate protein-protein interactions during mismatch recognition and strand removal; a ternary complex is formed between MutS, MutL, and the mismatched DNA, which activates the MutH endonuclease.
Pssm-ID: 340403 [Multi-domain] Cd Length: 188 Bit Score: 278.55 E-value: 1.78e-88
MutL_Trans_hPMS2_like: transducer domain, having a ribosomal S5 domain 2-like fold, found in ...
222-364
8.52e-67
MutL_Trans_hPMS2_like: transducer domain, having a ribosomal S5 domain 2-like fold, found in proteins similar to human PSM2 (hPSM2). hPSM2 belongs to the DNA mismatch repair (MutL/MLH1/PMS2) family. This transducer domain is homologous to the second domain of the DNA gyrase B subunit, which is known to be important in nucleotide hydrolysis and the transduction of structural signals from ATP-binding site to the DNA breakage/reunion regions of the enzymes. Included in this group are proteins similar to yeast PMS1. The yeast MLH1-PMS1 and the human MLH1-PMS2 heterodimers play a role in meiosis. hMLH1-hPMS2 also participates in the repair of all DNA mismatch repair (MMR) substrates. Cells lacking hPMS2 have a strong mutator phenotype and display microsatellite instability (MSI). Mutation in hPMS2 causes predisposition to HPNCC and Turcot syndrome.
Pssm-ID: 239566 [Multi-domain] Cd Length: 142 Bit Score: 219.06 E-value: 8.52e-67
MutL_Trans: transducer domain, having a ribosomal S5 domain 2-like fold, conserved in the ...
223-359
6.76e-37
MutL_Trans: transducer domain, having a ribosomal S5 domain 2-like fold, conserved in the C-terminal domain of DNA mismatch repair (MutL/MLH1/PMS2) family. This transducer domain is homologous to the second domain of the DNA gyrase B subunit, which is known to be important in nucleotide hydrolysis and the transduction of structural signals from ATP-binding site to the DNA breakage/reunion regions of the enzymes. Included in this group are proteins similar to human MLH1, hPMS2, hPMS1, hMLH3 and E. coli MutL, MLH1 forms heterodimers with PMS2, PMS1 and MLH3. These three complexes have distinct functions in meiosis. hMLH1-hPMS2 also participates in the repair of all DNA mismatch repair (MMR) substrates. Roles for hMLH1-hPMS1 or hMLH1-hMLH3 in MMR have not been established. Cells lacking either hMLH1 or hPMS2 have a strong mutator phenotype and display microsatellite instability (MSI). Mutation in hMLH1 causes predisposition to HNPCC, Muir-Torre syndrome and Turcot syndrome (HNPCC variant). Mutation in hPMS2 causes predisposition to HPNCC and Turcot syndrome. Mutation in hMLH1 accounts for a large fraction of HNPCC families. There is no convincing evidence to support hPMS1 having a role in HNPCC predisposition. It has been suggested that hMLH3 may be a low risk gene for colorectal cancer; however there is little evidence to support it having a role in classical HNPCC. It has been suggested that during initiation of DNA mismatch repair in E. coli, the mismatch recognition protein MutS recruits MutL in the presence of ATP. The MutS(ATP)-MutL ternary complex formed, then recruits the latent endonuclease MutH.
Pssm-ID: 238405 [Multi-domain] Cd Length: 122 Bit Score: 134.59 E-value: 6.76e-37
MutL C terminal dimerisation domain; MutL and MutS are key components of the DNA repair ...
678-832
9.92e-36
MutL C terminal dimerisation domain; MutL and MutS are key components of the DNA repair machinery that corrects replication errors. MutS recognises mispaired or unpaired bases in a DNA duplex and in the presence of ATP, recruits MutL to form a DNA signaling complex for repair. The N terminal region of MutL contains the ATPase domain and the C terminal is involved in dimerisation.
Pssm-ID: 214857 [Multi-domain] Cd Length: 140 Bit Score: 132.09 E-value: 9.92e-36
MutL C terminal dimerization domain; MutL and MutS are key components of the DNA repair ...
679-833
4.77e-28
MutL C terminal dimerization domain; MutL and MutS are key components of the DNA repair machinery that corrects replication errors. MutS recognizes mispaired or unpaired bases in a DNA duplex and in the presence of ATP, recruits MutL to form a DNA signaling complex for repair. The N terminal region of MutL contains the ATPase domain and the C terminal is involved in dimerization.
Pssm-ID: 430147 Cd Length: 145 Bit Score: 110.39 E-value: 4.77e-28
DNA mismatch repair protein, C-terminal domain; This family represents the C-terminal domain ...
229-359
8.25e-24
DNA mismatch repair protein, C-terminal domain; This family represents the C-terminal domain of the mutL/hexB/PMS1 family. This domain has a ribosomal S5 domain 2-like fold.
Pssm-ID: 426060 [Multi-domain] Cd Length: 117 Bit Score: 97.18 E-value: 8.25e-24
MutL_Trans: transducer domain, having a ribosomal S5 domain 2-like fold, conserved in the ...
223-344
2.19e-19
MutL_Trans: transducer domain, having a ribosomal S5 domain 2-like fold, conserved in the C-terminal domain of type II DNA topoisomerases (Topo II) and DNA mismatch repair (MutL/MLH1/PMS2) proteins. This transducer domain is homologous to the second domain of the DNA gyrase B subunit, which is known to be important in nucleotide hydrolysis and the transduction of structural signals from ATP-binding site to the DNA breakage/reunion regions of the enzymes. The GyrB dimerizes in response to ATP binding, and is homologous to the N-terminal half of eukaryotic Topo II and the ATPase fragment of MutL. Type II DNA topoisomerases catalyze the ATP-dependent transport of one DNA duplex through another, in the process generating transient double strand breaks via covalent attachments to both DNA strands at the 5' positions. Included in this group are proteins similar to human MLH1 and PMS2. MLH1 forms a heterodimer with PMS2 which functions in meiosis and in DNA mismatch repair (MMR). Cells lacking either hMLH1 or hPMS2 have a strong mutator phenotype and display microsatellite instability (MSI). Mutation in hMLH1 accounts for a large fraction of Lynch syndrome (HNPCC) families.
Pssm-ID: 238202 [Multi-domain] Cd Length: 107 Bit Score: 84.23 E-value: 2.19e-19
MutL_Trans_hPMS1_like: transducer domain, having a ribosomal S5 domain 2-like fold, found in ...
222-354
1.95e-11
MutL_Trans_hPMS1_like: transducer domain, having a ribosomal S5 domain 2-like fold, found in proteins similar to human PSM1 (hPSM1) and yeast MLH2. hPSM1 and yMLH2 are members of the DNA mismatch repair (MutL/MLH1/PMS2) family. This transducer domain is homologous to the second domain of the DNA gyrase B subunit, which is known to be important in nucleotide hydrolysis and the transduction of structural signals from ATP-binding site to the DNA breakage/reunion regions of the enzymes. PMS1 forms a heterodimer with MLH1. The MLH1-PMS1 complex functions in meiosis. Loss of yMLH2 results in a small but significant decrease in spore viability and a significant increase in gene conversion frequencies. A role for hMLH1-hPMS1 in DNA mismatch repair has not been established. Mutation in hMLH1 accounts for a large fraction of Lynch syndrome (HNPCC) families, however there is no convincing evidence to support hPMS1 having a role in HNPCC predisposition.
Pssm-ID: 239567 [Multi-domain] Cd Length: 132 Bit Score: 62.29 E-value: 1.95e-11
MutL_Trans_MLH1: transducer domain, having a ribosomal S5 domain 2-like fold, found in ...
222-348
1.18e-09
MutL_Trans_MLH1: transducer domain, having a ribosomal S5 domain 2-like fold, found in proteins similar to yeast and human MLH1 (MutL homologue 1). This transducer domain is homologous to the second domain of the DNA gyrase B subunit, which is known to be important in nucleotide hydrolysis and the transduction of structural signals from ATP-binding site to the DNA breakage/reunion regions of the enzymes. MLH1 forms heterodimers with PMS2, PMS1 and MLH3. These three complexes have distinct functions in meiosis. hMLH1-hPMS2 also participates in the repair of all DNA mismatch repair (MMR) substrates. Roles for hMLH1-hPMS1 or hMLH1-hMLH3 in MMR have not been established. Cells lacking hMLH1 have a strong mutator phenotype and display microsatellite instability (MSI). Mutation in hMLH1 causes predisposition to HNPCC, Muir-Torre syndrome and Turcot syndrome (HNPCC variant). Mutation in hMLH1 accounts for a large fraction of HNPCC families.
Pssm-ID: 239565 [Multi-domain] Cd Length: 127 Bit Score: 56.86 E-value: 1.18e-09
Histidine kinase-, DNA gyrase B-, and HSP90-like ATPase; This family represents, additionally, ...
35-151
3.68e-09
Histidine kinase-, DNA gyrase B-, and HSP90-like ATPase; This family represents, additionally, the structurally related ATPase domains of histidine kinase, DNA gyrase B and HSP90.
Pssm-ID: 433332 [Multi-domain] Cd Length: 135 Bit Score: 55.80 E-value: 3.68e-09
Histidine kinase-, DNA gyrase B-, and HSP90-like ATPase; This family represents the ...
31-170
1.01e-06
Histidine kinase-, DNA gyrase B-, and HSP90-like ATPase; This family represents the structurally related ATPase domains of histidine kinase, DNA gyrase B and HSP90.
Pssm-ID: 460579 [Multi-domain] Cd Length: 109 Bit Score: 48.13 E-value: 1.01e-06
MutL_Trans_MutL: transducer domain, having a ribosomal S5 domain 2-like fold, found in ...
259-343
8.49e-05
MutL_Trans_MutL: transducer domain, having a ribosomal S5 domain 2-like fold, found in proteins similar to Escherichia coli MutL. EcMutL belongs to the DNA mismatch repair (MutL/MLH1/PMS2) family. This transducer domain is homologous to the second domain of the DNA gyrase B subunit, which is known to be important in nucleotide hydrolysis and the transduction of structural signals from the ATP-binding site to the DNA breakage/reunion regions of the enzymes. It has been suggested that during initiation of DNA mismatch repair in E. coli, the mismatch recognition protein MutS recruits MutL in the presence of ATP. The MutS(ATP)-MutL ternary complex formed, then recruits the latent endonuclease MutH. Prokaryotic MutS and MutL are homodimers.
Pssm-ID: 239564 [Multi-domain] Cd Length: 123 Bit Score: 42.96 E-value: 8.49e-05
MutL_Trans_MLH3: transducer domain, having a ribosomal S5 domain 2-like fold, found in ...
255-355
4.55e-04
MutL_Trans_MLH3: transducer domain, having a ribosomal S5 domain 2-like fold, found in proteins similar to yeast and human MLH3 (MutL homologue 3). MLH3 belongs to the DNA mismatch repair (MutL/MLH1/PMS2) family. This transducer domain is homologous to the second domain of the DNA gyrase B subunit, which is known to be important in nucleotide hydrolysis and the transduction of structural signals from ATP-binding site to the DNA breakage/reunion regions of the enzymes. MLH1 forms heterodimers with MLH3. The MLH1-MLH3 complex plays a role in meiosis. A role for hMLH1-hMLH3 in DNA mismatch repair (MMR) has not been established. It has been suggested that hMLH3 may be a low risk gene for colorectal cancer; however there is little evidence to support it having a role in classical HNPCC.
Pssm-ID: 239568 [Multi-domain] Cd Length: 141 Bit Score: 41.15 E-value: 4.55e-04
Histidine kinase-like ATPase domain of two-component sensor histidine kinases similar to ...
35-78
2.87e-03
Histidine kinase-like ATPase domain of two-component sensor histidine kinases similar to Escherichia coli K-12 DpiB, DcuS, and Bacillus subtilis CitS, DctS, and YufL; This family includes histidine kinase-like ATPase domains of Escherichia coli K-12 DpiB and DcuS, and Bacillus subtilis CitS, DctS and MalK histidine kinases (HKs) all of which are two component transduction systems (TCSs). E. coli K-12 DpiB (also known as CitA) is the histidine kinase (HK) of DpiA-DpiB, a two-component signal transduction system (TCS) required for the expression of citrate-specific fermentation genes and genes involved in plasmid inheritance. E. coli K-12 DcuS (also known as YjdH) is the HK of DcuS-DcuR, a TCS that in the presence of the extracellular C4-dicarboxlates, activates the expression of the genes of anaerobic fumarate respiration and of aerobic C4-dicarboxylate uptake. CitS is the HK of Bacillus subtilis CitS-CitT, a TCS which regulates expression of CitM, the Mg-citrate transporter. Bacillus subtilis DctS forms a tripartite sensor unit (DctS/DctA/DctB) for sensing C4 dicarboxylates. Bacillus subtilis MalK (also known as YfuL) is the HK of MalK-MalR (YufL-YufM) a TCS which regulates the expression of the malate transporters MaeN (YufR) and YflS, and is essential for utilization of malate in minimal medium. Proteins having this DpiB-CitA-like HATPase domain generally have sensor domains such as Cache and PAS, and a histidine kinase A (HisKA)-like SpoOB-type, alpha-helical domain.
Pssm-ID: 340392 [Multi-domain] Cd Length: 104 Bit Score: 38.04 E-value: 2.87e-03
Histidine kinase-like ATPase domain of two-component sensor histidine kinases similar to ...
29-83
3.54e-03
Histidine kinase-like ATPase domain of two-component sensor histidine kinases similar to Escherichia coli BasS; This family includes the histidine kinase-like ATPase (HATPase) domains of various two-component sensor histidine kinase (HKs) similar to Escherichia coli BasS HK of the BasS-BasR two-component regulatory system (TCS). Proteins having this HATPase domain also contain a histidine kinase dimerization and phosphoacceptor domain (HisKA); some contain a HAMP sensory domain, while some an N-terminal two-component sensor kinase domain.
Pssm-ID: 340417 [Multi-domain] Cd Length: 113 Bit Score: 38.16 E-value: 3.54e-03
Histidine kinase-like ATPase domain of type IIB topoisomerase, Topo VI, subunit B; This family ...
34-78
6.61e-03
Histidine kinase-like ATPase domain of type IIB topoisomerase, Topo VI, subunit B; This family includes the histidine kinase-like ATPase (HATPase) domain of the B subunit of topoisomerase VI (Topo VIB). Topo VI is a heterotetrameric complex composed of two TopVIA and two TopVIB subunits and is categorized as a type II B DNA topoisomerase. It is found in archaea and also in plants. Type II enzymes cleave both strands of a DNA duplex and pass a second duplex through the resulting break in an ATP-dependent mechanism. DNA cleavage by Topo VI generates two-nucleotide 5'-protruding ends.
Pssm-ID: 340410 [Multi-domain] Cd Length: 203 Bit Score: 38.87 E-value: 6.61e-03
Histidine kinase-like ATPase domain of human microrchidia (MORC) family CW-type zinc finger ...
38-78
7.59e-03
Histidine kinase-like ATPase domain of human microrchidia (MORC) family CW-type zinc finger proteins MORC1-4, and related domains; This family includes the histidine kinase-like ATPase (HATPase) domain of human microrchidia (MORC) family CW-type zinc finger proteins MORC1-4, and related domains. In addition to the HATPase domain, MORC family proteins have a CW-type zinc finger domain containing four conserved cysteines and two conserved tryptophans, and coiled-coil domains at the carboxy-terminus. MORC1 has cross-species differential methylation in association with early life stress, and genome-wide association with major depressive disorder (MDD). MORC2 is involved in several nuclear processes, including transcription modulation and DNA damage repair, and exhibits a cytosolic function in lipogenesis, adipogenic differentiation, and lipid homeostasis by increasing the activity of ACLY. MORC3 regulates p53, and is an antiviral factor which plays an important role during HSV-1 and HCMV infection, and is a positive regulator of influenza virus transcription. MORC4 is highly expressed in a subset of diffuse large B-cell lymphomas and has potential as a lymphoma biomarker.
Pssm-ID: 340408 [Multi-domain] Cd Length: 118 Bit Score: 37.39 E-value: 7.59e-03
Database: CDSEARCH/cdd Low complexity filter: no Composition Based Adjustment: yes E-value threshold: 0.01
References:
Wang J et al. (2023), "The conserved domain database in 2023", Nucleic Acids Res.51(D)384-8.
Lu S et al. (2020), "The conserved domain database in 2020", Nucleic Acids Res.48(D)265-8.
Marchler-Bauer A et al. (2017), "CDD/SPARCLE: functional classification of proteins via subfamily domain architectures.", Nucleic Acids Res.45(D)200-3.
of the residues that compose this conserved feature have been mapped to the query sequence.
Click on the triangle to view details about the feature, including a multiple sequence alignment
of your query sequence and the protein sequences used to curate the domain model,
where hash marks (#) above the aligned sequences show the location of the conserved feature residues.
The thumbnail image, if present, provides an approximate view of the feature's location in 3 dimensions.
Click on the triangle for interactive 3D structure viewing options.
Functional characterization of the conserved domain architecture found on the query.
Click here to see more details.
This image shows a graphical summary of conserved domains identified on the query sequence.
The Show Concise/Full Display button at the top of the page can be used to select the desired level of detail: only top scoring hits
(labeled illustration) or all hits
(labeled illustration).
Domains are color coded according to superfamilies
to which they have been assigned. Hits with scores that pass a domain-specific threshold
(specific hits) are drawn in bright colors.
Others (non-specific hits) and
superfamily placeholders are drawn in pastel colors.
if a domain or superfamily has been annotated with functional sites (conserved features),
they are mapped to the query sequence and indicated through sets of triangles
with the same color and shade of the domain or superfamily that provides the annotation. Mouse over the colored bars or triangles to see descriptions of the domains and features.
click on the bars or triangles to view your query sequence embedded in a multiple sequence alignment of the proteins used to develop the corresponding domain model.
The table lists conserved domains identified on the query sequence. Click on the plus sign (+) on the left to display full descriptions, alignments, and scores.
Click on the domain model's accession number to view the multiple sequence alignment of the proteins used to develop the corresponding domain model.
To view your query sequence embedded in that multiple sequence alignment, click on the colored bars in the Graphical Summary portion of the search results page,
or click on the triangles, if present, that represent functional sites (conserved features)
mapped to the query sequence.
Concise Display shows only the best scoring domain model, in each hit category listed below except non-specific hits, for each region on the query sequence.
(labeled illustration) Standard Display shows only the best scoring domain model from each source, in each hit category listed below for each region on the query sequence.
(labeled illustration) Full Display shows all domain models, in each hit category below, that meet or exceed the RPS-BLAST threshold for statistical significance.
(labeled illustration) Four types of hits can be shown, as available,
for each region on the query sequence:
specific hits meet or exceed a domain-specific e-value threshold
(illustrated example)
and represent a very high confidence that the query sequence belongs to the same protein family as the sequences use to create the domain model
non-specific hits
meet or exceed the RPS-BLAST threshold for statistical significance (default E-value cutoff of 0.01, or an E-value selected by user via the
advanced search options)
the domain superfamily to which the specific and non-specific hits belong
multi-domain models that were computationally detected and are likely to contain multiple single domains
Retrieve proteins that contain one or more of the domains present in the query sequence, using the Conserved Domain Architecture Retrieval Tool
(CDART).
Modify your query to search against a different database and/or use advanced search options