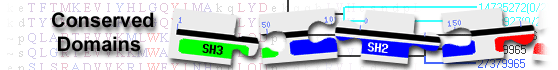Fatty acid synthase subunit alpha Acyl carrier domain; This is the acyl carrier domain (ACP) ...
142-302
2.99e-87
Fatty acid synthase subunit alpha Acyl carrier domain; This is the acyl carrier domain (ACP) found in fatty acid synthase subunit alpha (FAS2) EC:2.3.1.86.The fungal type I fatty acid synthase (FAS) is a 2.6 MDa multienzyme complex, catalyzing all necessary steps for the synthesis of long acyl chains. To be catalytically competent, the FAS must be activated by a posttranslational modification of the central acyl carrier domain (ACP) by an intrinsic phosphopantetheine transferase (PPT).
:
Pssm-ID: 465711 Cd Length: 162 Bit Score: 281.39 E-value: 2.99e-87
Fatty acid synthase type I helical domain; This domain is found in the fatty acid synthase ...
332-534
1.27e-76
Fatty acid synthase type I helical domain; This domain is found in the fatty acid synthase (FAS) complex present in species such as Mycobacterium smegmatis and Thermomyces lanuginosus. FAS is a homo-hexameric enzyme that catalyzes synthesis of fatty acid precursors of mycolic acids. This domain is composed of dimerization module 1 (DM1) and four-helix bundle (4HB), both of which are conserved parts of the acetyl transferase.
:
Pssm-ID: 465705 Cd Length: 203 Bit Score: 252.69 E-value: 1.27e-76
phosphopantetheine--protein transferase domain; This model models a domain active in ...
1765-1886
2.44e-41
phosphopantetheine--protein transferase domain; This model models a domain active in transferring the phophopantetheine prosthetic group to its attachment site on enzymes and carrier proteins. Many members of this family are small proteins that act on the acyl carrier protein involved in fatty acid biosynthesis. Some members are domains of larger proteins involved specialized pathways for the synthesis of unusual molecules including polyketides, atypical fatty acids, and antibiotics. [Protein fate, Protein modification and repair]
:
Pssm-ID: 273136 [Multi-domain] Cd Length: 128 Bit Score: 148.74 E-value: 2.44e-41
Malonyl CoA-acyl carrier protein transacylase [Lipid transport and metabolism]; Malonyl ...
16-73
4.69e-08
Malonyl CoA-acyl carrier protein transacylase [Lipid transport and metabolism]; Malonyl CoA-acyl carrier protein transacylase is part of the Pathway/BioSystem: Fatty acid biosynthesis
The actual alignment was detected with superfamily member COG0331:
Pssm-ID: 440100 [Multi-domain] Cd Length: 306 Bit Score: 56.67 E-value: 4.69e-08
ketoacyl reductase (KR) domain of fungal-type fatty acid synthase (fFAS), classical (c)-like SDRs; KR domain of fungal-type fatty acid synthase (FAS), type I. Fungal-type FAS is a heterododecameric FAS composed of alpha and beta multifunctional polypeptide chains. The KR, an SDR family member, is located centrally in the alpha chain. KR catalyzes the NADP-dependent reduction of ketoacyl-ACP to hydroxyacyl-ACP. KR shares the critical active site Tyr of the Classical SDR and has partial identity of the active site tetrad, but the upstream Asn is replaced in KR by Met. As in other SDRs, there is a glycine rich NAD-binding motif, but the pattern found in KR does not match the classical SDRs, and is not strictly conserved within this group. SDRs are a functionally diverse family of oxidoreductases that have a single domain with a structurally conserved Rossmann fold (alpha/beta folding pattern with a central beta-sheet), an NAD(P)(H)-binding region, and a structurally diverse C-terminal region. Classical SDRs are typically about 250 residues long, while extended SDRs are approximately 350 residues. Sequence identity between different SDR enzymes are typically in the 15-30% range, but the enzymes share the Rossmann fold NAD-binding motif and characteristic NAD-binding and catalytic sequence patterns. These enzymes catalyze a wide range of activities including the metabolism of steroids, cofactors, carbohydrates, lipids, aromatic compounds, and amino acids, and act in redox sensing. Classical SDRs have an TGXXX[AG]XG cofactor binding motif and a YXXXK active site motif, with the Tyr residue of the active site motif serving as a critical catalytic residue (Tyr-151, human prostaglandin dehydrogenase (PGDH) numbering). In addition to the Tyr and Lys, there is often an upstream Ser (Ser-138, PGDH numbering) and/or an Asn (Asn-107, PGDH numbering) contributing to the active site; while substrate binding is in the C-terminal region, which determines specificity. The standard reaction mechanism is a 4-pro-S hydride transfer and proton relay involving the conserved Tyr and Lys, a water molecule stabilized by Asn, and nicotinamide. Extended SDRs have additional elements in the C-terminal region, and typically have a TGXXGXXG cofactor binding motif. Complex (multidomain) SDRs such as ketoreductase domains of fatty acid synthase have a GGXGXXG NAD(P)-binding motif and an altered active site motif (YXXXN). Fungal type KRs have a TGXXXGX(1-2)G NAD(P)-binding motif. Some atypical SDRs have lost catalytic activity and/or have an unusual NAD(P)-binding motif and missing or unusual active site residues. Reactions catalyzed within the SDR family include isomerization, decarboxylation, epimerization, C=N bond reduction, dehydratase activity, dehalogenation, Enoyl-CoA reduction, and carbonyl-alcohol oxidoreduction.
Pssm-ID: 187653 [Multi-domain] Cd Length: 259 Bit Score: 508.27 E-value: 5.15e-167
Fatty acid synthase subunit alpha Acyl carrier domain; This is the acyl carrier domain (ACP) ...
142-302
2.99e-87
Fatty acid synthase subunit alpha Acyl carrier domain; This is the acyl carrier domain (ACP) found in fatty acid synthase subunit alpha (FAS2) EC:2.3.1.86.The fungal type I fatty acid synthase (FAS) is a 2.6 MDa multienzyme complex, catalyzing all necessary steps for the synthesis of long acyl chains. To be catalytically competent, the FAS must be activated by a posttranslational modification of the central acyl carrier domain (ACP) by an intrinsic phosphopantetheine transferase (PPT).
Pssm-ID: 465711 Cd Length: 162 Bit Score: 281.39 E-value: 2.99e-87
Fatty acid synthase type I helical domain; This domain is found in the fatty acid synthase ...
332-534
1.27e-76
Fatty acid synthase type I helical domain; This domain is found in the fatty acid synthase (FAS) complex present in species such as Mycobacterium smegmatis and Thermomyces lanuginosus. FAS is a homo-hexameric enzyme that catalyzes synthesis of fatty acid precursors of mycolic acids. This domain is composed of dimerization module 1 (DM1) and four-helix bundle (4HB), both of which are conserved parts of the acetyl transferase.
Pssm-ID: 465705 Cd Length: 203 Bit Score: 252.69 E-value: 1.27e-76
phosphopantetheine--protein transferase domain; This model models a domain active in ...
1765-1886
2.44e-41
phosphopantetheine--protein transferase domain; This model models a domain active in transferring the phophopantetheine prosthetic group to its attachment site on enzymes and carrier proteins. Many members of this family are small proteins that act on the acyl carrier protein involved in fatty acid biosynthesis. Some members are domains of larger proteins involved specialized pathways for the synthesis of unusual molecules including polyketides, atypical fatty acids, and antibiotics. [Protein fate, Protein modification and repair]
Pssm-ID: 273136 [Multi-domain] Cd Length: 128 Bit Score: 148.74 E-value: 2.44e-41
4'-phosphopantetheinyl transferase superfamily; Members of this family transfers the 4 ...
1768-1864
5.72e-17
4'-phosphopantetheinyl transferase superfamily; Members of this family transfers the 4'-phosphopantetheine (4'-PP) moiety from coenzyme A (CoA) to the invariant serine of pfam00550. This post-translational modification renders holo-ACP capable of acyl group activation via thioesterification of the cysteamine thiol of 4'-PP. This superfamily consists of two subtypes: The ACPS type and the Sfp type. The structure of the Sfp type is known, which shows the active site accommodates a magnesium ion. The most highly conserved regions of the alignment are involved in binding the magnesium ion.
Pssm-ID: 426364 [Multi-domain] Cd Length: 111 Bit Score: 78.42 E-value: 5.72e-17
Beta-ketoacyl synthase, N-terminal domain; The structure of beta-ketoacyl synthase is similar ...
1182-1383
1.18e-12
Beta-ketoacyl synthase, N-terminal domain; The structure of beta-ketoacyl synthase is similar to that of the thiolase family (pfam00108) and also chalcone synthase. The active site of beta-ketoacyl synthase is located between the N and C-terminal domains. The N-terminal domain contains most of the structures involved in dimer formation and also the active site cysteine.
Pssm-ID: 425468 [Multi-domain] Cd Length: 251 Bit Score: 69.97 E-value: 1.18e-12
Malonyl CoA-acyl carrier protein transacylase [Lipid transport and metabolism]; Malonyl ...
16-73
4.69e-08
Malonyl CoA-acyl carrier protein transacylase [Lipid transport and metabolism]; Malonyl CoA-acyl carrier protein transacylase is part of the Pathway/BioSystem: Fatty acid biosynthesis
Pssm-ID: 440100 [Multi-domain] Cd Length: 306 Bit Score: 56.67 E-value: 4.69e-08
ketoacyl reductase (KR) domain of fungal-type fatty acid synthase (fFAS), classical (c)-like SDRs; KR domain of fungal-type fatty acid synthase (FAS), type I. Fungal-type FAS is a heterododecameric FAS composed of alpha and beta multifunctional polypeptide chains. The KR, an SDR family member, is located centrally in the alpha chain. KR catalyzes the NADP-dependent reduction of ketoacyl-ACP to hydroxyacyl-ACP. KR shares the critical active site Tyr of the Classical SDR and has partial identity of the active site tetrad, but the upstream Asn is replaced in KR by Met. As in other SDRs, there is a glycine rich NAD-binding motif, but the pattern found in KR does not match the classical SDRs, and is not strictly conserved within this group. SDRs are a functionally diverse family of oxidoreductases that have a single domain with a structurally conserved Rossmann fold (alpha/beta folding pattern with a central beta-sheet), an NAD(P)(H)-binding region, and a structurally diverse C-terminal region. Classical SDRs are typically about 250 residues long, while extended SDRs are approximately 350 residues. Sequence identity between different SDR enzymes are typically in the 15-30% range, but the enzymes share the Rossmann fold NAD-binding motif and characteristic NAD-binding and catalytic sequence patterns. These enzymes catalyze a wide range of activities including the metabolism of steroids, cofactors, carbohydrates, lipids, aromatic compounds, and amino acids, and act in redox sensing. Classical SDRs have an TGXXX[AG]XG cofactor binding motif and a YXXXK active site motif, with the Tyr residue of the active site motif serving as a critical catalytic residue (Tyr-151, human prostaglandin dehydrogenase (PGDH) numbering). In addition to the Tyr and Lys, there is often an upstream Ser (Ser-138, PGDH numbering) and/or an Asn (Asn-107, PGDH numbering) contributing to the active site; while substrate binding is in the C-terminal region, which determines specificity. The standard reaction mechanism is a 4-pro-S hydride transfer and proton relay involving the conserved Tyr and Lys, a water molecule stabilized by Asn, and nicotinamide. Extended SDRs have additional elements in the C-terminal region, and typically have a TGXXGXXG cofactor binding motif. Complex (multidomain) SDRs such as ketoreductase domains of fatty acid synthase have a GGXGXXG NAD(P)-binding motif and an altered active site motif (YXXXN). Fungal type KRs have a TGXXXGX(1-2)G NAD(P)-binding motif. Some atypical SDRs have lost catalytic activity and/or have an unusual NAD(P)-binding motif and missing or unusual active site residues. Reactions catalyzed within the SDR family include isomerization, decarboxylation, epimerization, C=N bond reduction, dehydratase activity, dehalogenation, Enoyl-CoA reduction, and carbonyl-alcohol oxidoreduction.
Pssm-ID: 187653 [Multi-domain] Cd Length: 259 Bit Score: 508.27 E-value: 5.15e-167
ketoacyl reductase (KR) domain of fungal-type fatty acid synthase (fFAS)-like, classical (c)-like SDRs; KR domain of FAS, including the fungal-type multidomain FAS alpha chain, and the single domain daunorubicin C-13 ketoreductase. Fungal-type FAS is a heterododecameric FAS composed of alpha and beta multifunctional polypeptide chains. The KR, an SDR family member is located centrally in the alpha chain. KR catalyzes the NADP-dependent reduction of ketoacyl-ACP to hydroxyacyl-ACP. KR shares the critical active site Tyr of the classical SDR and has partial identity of the active site tetrad, but the upstream Asn is replaced in KR by Met. As in other SDRs, there is a glycine rich NAD(P)-binding motif, but the pattern found in KR does not match the classical SDRs, and is not strictly conserved within this group. Daunorubicin is a clinically important therapeutic compound used in some cancer treatments. Single domain daunorubicin C-13 ketoreductase is member of the classical SDR family with a canonical glycine-rich NAD(P)-binding motif, but lacking a complete match to the active site tetrad characteristic of this group. The critical Tyr, plus the Lys and upstream Asn are present, but the catalytic Ser is replaced, generally by Gln. SDRs are a functionally diverse family of oxidoreductases that have a single domain with a structurally conserved Rossmann fold (alpha/beta folding pattern with a central beta-sheet), an NAD(P)(H)-binding region, and a structurally diverse C-terminal region. Classical SDRs are typically about 250 residues long, while extended SDRs are approximately 350 residues. Sequence identity between different SDR enzymes are typically in the 15-30% range, but the enzymes share the Rossmann fold NAD-binding motif and characteristic NAD-binding and catalytic sequence patterns. These enzymes catalyze a wide range of activities including the metabolism of steroids, cofactors, carbohydrates, lipids, aromatic compounds, and amino acids, and act in redox sensing. Classical SDRs have an TGXXX[AG]XG cofactor binding motif and a YXXXK active site motif, with the Tyr residue of the active site motif serving as a critical catalytic residue (Tyr-151, human prostaglandin dehydrogenase (PGDH) numbering). In addition to the Tyr and Lys, there is often an upstream Ser (Ser-138, PGDH numbering) and/or an Asn (Asn-107, PGDH numbering) contributing to the active site; while substrate binding is in the C-terminal region, which determines specificity. The standard reaction mechanism is a 4-pro-S hydride transfer and proton relay involving the conserved Tyr and Lys, a water molecule stabilized by Asn, and nicotinamide. Extended SDRs have additional elements in the C-terminal region, and typically have a TGXXGXXG cofactor binding motif. Complex (multidomain) SDRs such as ketoreductase domains of fatty acid synthase have a GGXGXXG NAD(P)-binding motif and an altered active site motif (YXXXN). Fungal type KRs have a TGXXXGX(1-2)G NAD(P)-binding motif. Some atypical SDRs have lost catalytic activity and/or have an unusual NAD(P)-binding motif and missing or unusual active site residues. Reactions catalyzed within the SDR family include isomerization, decarboxylation, epimerization, C=N bond reduction, dehydratase activity, dehalogenation, Enoyl-CoA reduction, and carbonyl-alcohol oxidoreduction.
Pssm-ID: 187633 [Multi-domain] Cd Length: 248 Bit Score: 438.65 E-value: 1.46e-141
"elongating" condensing enzymes are a subclass of decarboxylating condensing enzymes, ...
1135-1655
3.45e-123
"elongating" condensing enzymes are a subclass of decarboxylating condensing enzymes, including beta-ketoacyl [ACP] synthase, type I and II and polyketide synthases.They are characterized by the utlization of acyl carrier protein (ACP) thioesters as primer substrates, as well as the nature of their active site residues.
Pssm-ID: 238424 [Multi-domain] Cd Length: 407 Bit Score: 394.11 E-value: 3.45e-123
Fatty acid synthase subunit alpha Acyl carrier domain; This is the acyl carrier domain (ACP) ...
142-302
2.99e-87
Fatty acid synthase subunit alpha Acyl carrier domain; This is the acyl carrier domain (ACP) found in fatty acid synthase subunit alpha (FAS2) EC:2.3.1.86.The fungal type I fatty acid synthase (FAS) is a 2.6 MDa multienzyme complex, catalyzing all necessary steps for the synthesis of long acyl chains. To be catalytically competent, the FAS must be activated by a posttranslational modification of the central acyl carrier domain (ACP) by an intrinsic phosphopantetheine transferase (PPT).
Pssm-ID: 465711 Cd Length: 162 Bit Score: 281.39 E-value: 2.99e-87
Fatty acid synthase type I helical domain; This domain is found in the fatty acid synthase ...
332-534
1.27e-76
Fatty acid synthase type I helical domain; This domain is found in the fatty acid synthase (FAS) complex present in species such as Mycobacterium smegmatis and Thermomyces lanuginosus. FAS is a homo-hexameric enzyme that catalyzes synthesis of fatty acid precursors of mycolic acids. This domain is composed of dimerization module 1 (DM1) and four-helix bundle (4HB), both of which are conserved parts of the acetyl transferase.
Pssm-ID: 465705 Cd Length: 203 Bit Score: 252.69 E-value: 1.27e-76
Short-chain dehydrogenases/reductases (SDR); SDRs are a functionally diverse family of ...
678-917
3.90e-44
Short-chain dehydrogenases/reductases (SDR); SDRs are a functionally diverse family of oxidoreductases that have a single domain with a structurally conserved Rossmann fold (alpha/beta folding pattern with a central beta-sheet), an NAD(P)(H)-binding region, and a structurally diverse C-terminal region. Classical SDRs are typically about 250 residues long, while extended SDRs are approximately 350 residues. Sequence identity between different SDR enzymes are typically in the 15-30% range, but the enzymes share the Rossmann fold NAD-binding motif and characteristic NAD-binding and catalytic sequence patterns. These enzymes catalyze a wide range of activities including the metabolism of steroids, cofactors, carbohydrates, lipids, aromatic compounds, and amino acids, and act in redox sensing. Classical SDRs have an TGXXX[AG]XG cofactor binding motif and a YXXXK active site motif, with the Tyr residue of the active site motif serving as a critical catalytic residue (Tyr-151, human prostaglandin dehydrogenase (PGDH) numbering). In addition to the Tyr and Lys, there is often an upstream Ser (Ser-138, PGDH numbering) and/or an Asn (Asn-107, PGDH numbering) contributing to the active site; while substrate binding is in the C-terminal region, which determines specificity. The standard reaction mechanism is a 4-pro-S hydride transfer and proton relay involving the conserved Tyr and Lys, a water molecule stabilized by Asn, and nicotinamide. Extended SDRs have additional elements in the C-terminal region, and typically have a TGXXGXXG cofactor binding motif. Complex (multidomain) SDRs such as ketoreductase (KR) domains of fatty acid synthase have a GGXGXXG NAD(P)-binding motif and an altered active site motif (YXXXN). Fungal type KRs have a TGXXXGX(1-2)G NAD(P)-binding motif. Some atypical SDRs have lost catalytic activity and/or have an unusual NAD(P)-binding motif and missing or unusual active site residues. Reactions catalyzed within the SDR family include isomerization, decarboxylation, epimerization, C=N bond reduction, dehydratase activity, dehalogenation, Enoyl-CoA reduction, and carbonyl-alcohol oxidoreduction.
Pssm-ID: 187535 [Multi-domain] Cd Length: 186 Bit Score: 158.83 E-value: 3.90e-44
phosphopantetheine--protein transferase domain; This model models a domain active in ...
1765-1886
2.44e-41
phosphopantetheine--protein transferase domain; This model models a domain active in transferring the phophopantetheine prosthetic group to its attachment site on enzymes and carrier proteins. Many members of this family are small proteins that act on the acyl carrier protein involved in fatty acid biosynthesis. Some members are domains of larger proteins involved specialized pathways for the synthesis of unusual molecules including polyketides, atypical fatty acids, and antibiotics. [Protein fate, Protein modification and repair]
Pssm-ID: 273136 [Multi-domain] Cd Length: 128 Bit Score: 148.74 E-value: 2.44e-41
3-oxoacyl-(acyl-carrier-protein) synthase [Lipid transport and metabolism, Secondary ...
1180-1647
2.71e-26
3-oxoacyl-(acyl-carrier-protein) synthase [Lipid transport and metabolism, Secondary metabolites biosynthesis, transport and catabolism]; 3-oxoacyl-(acyl-carrier-protein) synthase is part of the Pathway/BioSystem: Fatty acid biosynthesis
Pssm-ID: 440073 [Multi-domain] Cd Length: 409 Bit Score: 113.65 E-value: 2.71e-26
Beta-ketoacyl-acyl carrier protein (ACP) synthase (KAS), type I and II. KASs are responsible ...
1185-1647
3.89e-26
Beta-ketoacyl-acyl carrier protein (ACP) synthase (KAS), type I and II. KASs are responsible for the elongation steps in fatty acid biosynthesis. KASIII catalyses the initial condensation and KAS I and II catalyze further elongation steps by Claisen condensation of malonyl-acyl carrier protein (ACP) with acyl-ACP.
Pssm-ID: 238430 [Multi-domain] Cd Length: 406 Bit Score: 113.40 E-value: 3.89e-26
4'-phosphopantetheinyl transferase superfamily; Members of this family transfers the 4 ...
1768-1864
5.72e-17
4'-phosphopantetheinyl transferase superfamily; Members of this family transfers the 4'-phosphopantetheine (4'-PP) moiety from coenzyme A (CoA) to the invariant serine of pfam00550. This post-translational modification renders holo-ACP capable of acyl group activation via thioesterification of the cysteamine thiol of 4'-PP. This superfamily consists of two subtypes: The ACPS type and the Sfp type. The structure of the Sfp type is known, which shows the active site accommodates a magnesium ion. The most highly conserved regions of the alignment are involved in binding the magnesium ion.
Pssm-ID: 426364 [Multi-domain] Cd Length: 111 Bit Score: 78.42 E-value: 5.72e-17
Beta-ketoacyl synthase, N-terminal domain; The structure of beta-ketoacyl synthase is similar ...
1182-1383
1.18e-12
Beta-ketoacyl synthase, N-terminal domain; The structure of beta-ketoacyl synthase is similar to that of the thiolase family (pfam00108) and also chalcone synthase. The active site of beta-ketoacyl synthase is located between the N and C-terminal domains. The N-terminal domain contains most of the structures involved in dimer formation and also the active site cysteine.
Pssm-ID: 425468 [Multi-domain] Cd Length: 251 Bit Score: 69.97 E-value: 1.18e-12
Beta-ketoacyl synthase, C-terminal domain; The structure of beta-ketoacyl synthase is similar ...
1522-1612
1.35e-08
Beta-ketoacyl synthase, C-terminal domain; The structure of beta-ketoacyl synthase is similar to that of the thiolase family (pfam00108) and also chalcone synthase. The active site of beta-ketoacyl synthase is located between the N and C-terminal domains.
Pssm-ID: 426989 Cd Length: 118 Bit Score: 54.50 E-value: 1.35e-08
NAD(P)-dependent dehydrogenase, short-chain alcohol dehydrogenase family [Lipid transport and ...
673-895
4.35e-08
NAD(P)-dependent dehydrogenase, short-chain alcohol dehydrogenase family [Lipid transport and metabolism]; NAD(P)-dependent dehydrogenase, short-chain alcohol dehydrogenase family is part of the Pathway/BioSystem: Fatty acid biosynthesis
Pssm-ID: 440651 [Multi-domain] Cd Length: 249 Bit Score: 56.33 E-value: 4.35e-08
Malonyl CoA-acyl carrier protein transacylase [Lipid transport and metabolism]; Malonyl ...
16-73
4.69e-08
Malonyl CoA-acyl carrier protein transacylase [Lipid transport and metabolism]; Malonyl CoA-acyl carrier protein transacylase is part of the Pathway/BioSystem: Fatty acid biosynthesis
Pssm-ID: 440100 [Multi-domain] Cd Length: 306 Bit Score: 56.67 E-value: 4.69e-08
holo-[acyl-carrier-protein] synthase; Formerly dpj. This enzyme adds the prosthetic group, ...
1768-1883
1.28e-07
holo-[acyl-carrier-protein] synthase; Formerly dpj. This enzyme adds the prosthetic group, phosphopantethiene, to the acyl carrier protein (ACP) apo-enzyme to generate the holo-enzyme. Related phosphopantethiene--protein transferases also exist. There is an orthologous domain in eukaryotic proteins. [Fatty acid and phospholipid metabolism, Biosynthesis]
Pssm-ID: 273114 [Multi-domain] Cd Length: 121 Bit Score: 52.00 E-value: 1.28e-07
RhlG and related beta-ketoacyl reductases, classical (c) SDRs; Pseudomonas aeruginosa RhlG is ...
674-903
8.11e-07
RhlG and related beta-ketoacyl reductases, classical (c) SDRs; Pseudomonas aeruginosa RhlG is an SDR-family beta-ketoacyl reductase involved in Rhamnolipid biosynthesis. RhlG is similar to but distinct from the FabG family of beta-ketoacyl-acyl carrier protein (ACP) of type II fatty acid synthesis. RhlG and related proteins are classical SDRs, with a canonical active site tetrad and glycine-rich NAD(P)-binding motif. SDRs are a functionally diverse family of oxidoreductases that have a single domain with a structurally conserved Rossmann fold (alpha/beta folding pattern with a central beta-sheet), an NAD(P)(H)-binding region, and a structurally diverse C-terminal region. Classical SDRs are typically about 250 residues long, while extended SDRs are approximately 350 residues. Sequence identity between different SDR enzymes are typically in the 15-30% range, but the enzymes share the Rossmann fold NAD-binding motif and characteristic NAD-binding and catalytic sequence patterns. These enzymes catalyze a wide range of activities including the metabolism of steroids, cofactors, carbohydrates, lipids, aromatic compounds, and amino acids, and act in redox sensing. Classical SDRs have an TGXXX[AG]XG cofactor binding motif and a YXXXK active site motif, with the Tyr residue of the active site motif serving as a critical catalytic residue (Tyr-151, human 15-hydroxyprostaglandin dehydrogenase (15-PGDH) numbering). In addition to the Tyr and Lys, there is often an upstream Ser (Ser-138, 15-PGDH numbering) and/or an Asn (Asn-107, 15-PGDH numbering) contributing to the active site; while substrate binding is in the C-terminal region, which determines specificity. The standard reaction mechanism is a 4-pro-S hydride transfer and proton relay involving the conserved Tyr and Lys, a water molecule stabilized by Asn, and nicotinamide. Extended SDRs have additional elements in the C-terminal region, and typically have a TGXXGXXG cofactor binding motif. Complex (multidomain) SDRs such as ketoreductase domains of fatty acid synthase have a GGXGXXG NAD(P)-binding motif and an altered active site motif (YXXXN). Fungal type ketoacyl reductases have a TGXXXGX(1-2)G NAD(P)-binding motif. Some atypical SDRs have lost catalytic activity and/or have an unusual NAD(P)-binding motif and missing or unusual active site residues. Reactions catalyzed within the SDR family include isomerization, decarboxylation, epimerization, C=N bond reduction, dehydratase activity, dehalogenation, Enoyl-CoA reduction, and carbonyl-alcohol oxidoreduction.
Pssm-ID: 187646 [Multi-domain] Cd Length: 250 Bit Score: 52.48 E-value: 8.11e-07
decarboxylating condensing enzymes; Family of enzymes that catalyze the formation of a new ...
1211-1431
9.22e-06
decarboxylating condensing enzymes; Family of enzymes that catalyze the formation of a new carbon-carbon bond by a decarboxylating Claisen-like condensation reaction. Members are involved in the synthesis of fatty acids and polyketides, a diverse group of natural products. Both pathways are an iterative series of additions of small carbon units, usually acetate, to a nascent acyl group. There are 2 classes of decarboxylating condensing enzymes, which can be distinguished by sequence similarity, type of active site residues and type of primer units (acetyl CoA or acyl carrier protein (ACP) linked units).
Pssm-ID: 238421 [Multi-domain] Cd Length: 332 Bit Score: 49.94 E-value: 9.22e-06
classical (c) SDRs; SDRs are a functionally diverse family of oxidoreductases that have a ...
678-862
1.04e-05
classical (c) SDRs; SDRs are a functionally diverse family of oxidoreductases that have a single domain with a structurally conserved Rossmann fold (alpha/beta folding pattern with a central beta-sheet), an NAD(P)(H)-binding region, and a structurally diverse C-terminal region. Classical SDRs are typically about 250 residues long, while extended SDRs are approximately 350 residues. Sequence identity between different SDR enzymes are typically in the 15-30% range, but the enzymes share the Rossmann fold NAD-binding motif and characteristic NAD-binding and catalytic sequence patterns. These enzymes catalyze a wide range of activities including the metabolism of steroids, cofactors, carbohydrates, lipids, aromatic compounds, and amino acids, and act in redox sensing. Classical SDRs have an TGXXX[AG]XG cofactor binding motif and a YXXXK active site motif, with the Tyr residue of the active site motif serving as a critical catalytic residue (Tyr-151, human prostaglandin dehydrogenase (PGDH) numbering). In addition to the Tyr and Lys, there is often an upstream Ser (Ser-138, PGDH numbering) and/or an Asn (Asn-107, PGDH numbering) contributing to the active site; while substrate binding is in the C-terminal region, which determines specificity. The standard reaction mechanism is a 4-pro-S hydride transfer and proton relay involving the conserved Tyr and Lys, a water molecule stabilized by Asn, and nicotinamide. Extended SDRs have additional elements in the C-terminal region, and typically have a TGXXGXXG cofactor binding motif. Complex (multidomain) SDRs such as ketoreductase domains of fatty acid synthase have a GGXGXXG NAD(P)-binding motif and an altered active site motif (YXXXN). Fungal type ketoacyl reductases have a TGXXXGX(1-2)G NAD(P)-binding motif. Some atypical SDRs have lost catalytic activity and/or have an unusual NAD(P)-binding motif and missing or unusual active site residues. Reactions catalyzed within the SDR family include isomerization, decarboxylation, epimerization, C=N bond reduction, dehydratase activity, dehalogenation, Enoyl-CoA reduction, and carbonyl-alcohol oxidoreduction.
Pssm-ID: 212491 [Multi-domain] Cd Length: 234 Bit Score: 48.82 E-value: 1.04e-05
mannitol dehydrogenase (MDH)-like, classical (c) SDRs; NADP-mannitol dehydrogenase catalyzes the conversion of fructose to mannitol, an acyclic 6-carbon sugar. MDH is a tetrameric member of the SDR family. This subgroup also includes various other tetrameric SDRs, including Pichia stipitis D-arabinitol dehydrogenase (aka polyol dehydrogenase), Candida albicans Sou1p, a sorbose reductase, and Candida parapsilosis (S)-specific carbonyl reductase (SCR, aka S-specific alcohol dehydrogenase) which catalyzes the enantioselective reduction of 2-hydroxyacetophenone into (S)-1-phenyl-1,2-ethanediol. SDRs are a functionally diverse family of oxidoreductases that have a single domain with a structurally conserved Rossmann fold (alpha/beta folding pattern with a central beta-sheet), an NAD(P)(H)-binding region, and a structurally diverse C-terminal region. Classical SDRs are typically about 250 residues long, while extended SDRS are approximately 350 residues. Sequence identity between different SDR enzymes are typically in the 15-30% range, but the enzymes share the Rossmann fold NAD-binding motif and characteristic NAD-binding and catalytic sequence patterns. These enzymes have a 3-glycine N-terminal NAD(P)(H)-binding pattern (typically, TGxxxGxG in classical SDRs and TGxxGxxG in extended SDRs), while substrate binding is in the C-terminal region. A critical catalytic Tyr residue (Tyr-151, human 15-hydroxyprostaglandin dehydrogenase (15-PGDH) numbering), is often found in a conserved YXXXK pattern. In addition to the Tyr and Lys, there is often an upstream Ser (Ser-138, 15-PGDH numbering) and/or an Asn (Asn-107, 15-PGDH numbering) or additional Ser, contributing to the active site. Substrates for these enzymes include sugars, steroids, alcohols, and aromatic compounds. The standard reaction mechanism is a proton relay involving the conserved Tyr and Lys, as well as Asn (or Ser).
Pssm-ID: 187610 [Multi-domain] Cd Length: 252 Bit Score: 47.71 E-value: 3.31e-05
beta-Keto acyl carrier protein reductase (BKR), involved in Type II FAS, classical (c) SDRs; ...
676-754
1.04e-04
beta-Keto acyl carrier protein reductase (BKR), involved in Type II FAS, classical (c) SDRs; This subgroup includes the Escherichai coli K12 BKR, FabG. BKR catalyzes the NADPH-dependent reduction of ACP in the first reductive step of de novo fatty acid synthesis (FAS). FAS consists of four elongation steps, which are repeated to extend the fatty acid chain through the addition of two-carbo units from malonyl acyl-carrier protein (ACP): condensation, reduction, dehydration, and a final reduction. Type II FAS, typical of plants and many bacteria, maintains these activities on discrete polypeptides, while type I FAS utilizes one or two multifunctional polypeptides. BKR resembles enoyl reductase, which catalyzes the second reduction step in FAS. SDRs are a functionally diverse family of oxidoreductases that have a single domain with structurally conserved Rossmann fold (alpha/beta folding pattern with a central beta-sheet) NAD(P)(H) binding region and a structurally diverse C-terminal region. Classical SDRs are typically about 250 residues long, while extended SDRS are approximately 350 residues. Sequence identity between different SDR enzymes are typically in the 15-30% range, but the enzymes share the Rossmann fold NAD binding motif and characteristic NAD-binding and catalytic sequence patterns. These enzymes have a 3-glycine N-terminal NAD(P)(H) binding pattern: TGxxxGxG in classical SDRs. Extended SDRs have additional elements in the C-terminal region, and typically have a TGXXGXXG cofactor binding motif. Complex (multidomain) SDRs such as ketoreductase domains of fatty acid synthase have a GGXGXXG NAD(P) binding motif and an altered active site motif (YXXXN). Fungal type type ketoacyl reductases have a TGXXXGX(1-2)G NAD(P)-binding motif. Some atypical SDRs have lost catalytic activity and/or have an unusual NAD(P) binding motif and missing or unusual active site residues. Reactions catalyzed within the SDR family include isomerization, decarboxylation, epimerization, C=N bond reduction, dehydratase activity, dehalogenation, Enoyl-CoA reduction, and carbonyl-alcohol oxidoreduction. A critical catalytic Tyr residue (Tyr-151, human 15-hydroxyprostaglandin dehydrogenase (15-PGDH) numbering), is often found in a conserved YXXXK pattern. In addition to the Tyr and Lys, there is often an upstream Ser (Ser-138, 15-PGDH numbering) and/or an Asn (Asn-107, 15-PGDH numbering) or additional Ser, contributing to the active site. Substrates for these enzymes include sugars, steroids, alcohols, and aromatic compounds. The standard reaction mechanism is a proton relay involving the conserved Tyr-151 and Lys-155, and well as Asn-111 (or Ser). Some SDR family members, including 17 beta-hydroxysteroid dehydrogenase contain an additional helix-turn-helix motif that is not generally found among SDRs.
Pssm-ID: 187594 [Multi-domain] Cd Length: 240 Bit Score: 46.00 E-value: 1.04e-04
putative beta-ketoacyl acyl carrier protein [ACP]reductase (BKR), subgroup 2, classical (c) SDR; This subgroup includes Rhizobium sp. NGR234 FabG1. The Escherichai coli K12 BKR, FabG, belongs to a different subgroup. BKR catalyzes the NADPH-dependent reduction of ACP in the first reductive step of de novo fatty acid synthesis (FAS). FAS consists of four elongation steps, which are repeated to extend the fatty acid chain through the addition of two-carbo units from malonyl acyl-carrier protein (ACP): condensation, reduction, dehydration, and a final reduction. Type II FAS, typical of plants and many bacteria, maintains these activities on discrete polypeptides, while type I FAS utilizes one or two multifunctional polypeptides. BKR resembles enoyl reductase, which catalyzes the second reduction step in FAS. SDRs are a functionally diverse family of oxidoreductases that have a single domain with a structurally conserved Rossmann fold (alpha/beta folding pattern with a central beta-sheet), an NAD(P)(H)-binding region, and a structurally diverse C-terminal region. Classical SDRs are typically about 250 residues long, while extended SDRS are approximately 350 residues. Sequence identity between different SDR enzymes are typically in the 15-30% range, but the enzymes share the Rossmann fold NAD-binding motif and characteristic NAD-binding and catalytic sequence patterns. These enzymes have a 3-glycine N-terminal NAD(P)(H)-binding pattern (typically, TGxxxGxG in classical SDRs and TGxxGxxG in extended SDRs), while substrate binding is in the C-terminal region. A critical catalytic Tyr residue (Tyr-151, human 15-hydroxyprostaglandin dehydrogenase (15-PGDH) numbering), is often found in a conserved YXXXK pattern. In addition to the Tyr and Lys, there is often an upstream Ser (Ser-138, 15-PGDH numbering) and/or an Asn (Asn-107, 15-PGDH numbering) or additional Ser, contributing to the active site. Substrates for these enzymes include sugars, steroids, alcohols, and aromatic compounds. The standard reaction mechanism is a proton relay involving the conserved Tyr and Lys, as well as Asn (or Ser). Some SDR family members, including 17 beta-hydroxysteroid dehydrogenase contain an additional helix-turn-helix motif that is not generally found among SDRs.
Pssm-ID: 187607 [Multi-domain] Cd Length: 246 Bit Score: 45.91 E-value: 1.11e-04
carbonyl reductase sniffer-like, classical (c) SDRs; Sniffer is an NADPH-dependent carbonyl ...
678-846
2.71e-03
carbonyl reductase sniffer-like, classical (c) SDRs; Sniffer is an NADPH-dependent carbonyl reductase of the classical SDR family. Studies in Drosophila melanogaster implicate Sniffer in the prevention of neurodegeneration due to aging and oxidative-stress. This subgroup also includes Rhodococcus sp. AD45 IsoH, which is an NAD-dependent 1-hydroxy-2-glutathionyl-2-methyl-3-butene dehydrogenase involved in isoprene metabolism, Aspergillus nidulans StcE encoded by a gene which is part of a proposed sterigmatocystin biosynthesis gene cluster, Bacillus circulans SANK 72073 BtrF encoded by a gene found in the butirosin biosynthesis gene cluster, and Aspergillus parasiticus nor-1 involved in the biosynthesis of aflatoxins. SDRs are a functionally diverse family of oxidoreductases that have a single domain with a structurally conserved Rossmann fold (alpha/beta folding pattern with a central beta-sheet), an NAD(P)(H)-binding region, and a structurally diverse C-terminal region. Classical SDRs are typically about 250 residues long, while extended SDRs are approximately 350 residues. Sequence identity between different SDR enzymes are typically in the 15-30% range, but the enzymes share the Rossmann fold NAD-binding motif and characteristic NAD-binding and catalytic sequence patterns. These enzymes catalyze a wide range of activities including the metabolism of steroids, cofactors, carbohydrates, lipids, aromatic compounds, and amino acids, and act in redox sensing. Classical SDRs have an TGXXX[AG]XG cofactor binding motif and a YXXXK active site motif, with the Tyr residue of the active site motif serving as a critical catalytic residue (Tyr-151, human 15-hydroxyprostaglandin dehydrogenase (15-PGDH) numbering). In addition to the Tyr and Lys, there is often an upstream Ser (Ser-138, 15-PGDH numbering) and/or an Asn (Asn-107, 15-PGDH numbering) contributing to the active site; while substrate binding is in the C-terminal region, which determines specificity. The standard reaction mechanism is a 4-pro-S hydride transfer and proton relay involving the conserved Tyr and Lys, a water molecule stabilized by Asn, and nicotinamide. Extended SDRs have additional elements in the C-terminal region, and typically have a TGXXGXXG cofactor binding motif. Complex (multidomain) SDRs such as ketoreductase domains of fatty acid synthase have a GGXGXXG NAD(P)-binding motif and an altered active site motif (YXXXN). Fungal type ketoacyl reductases have a TGXXXGX(1-2)G NAD(P)-binding motif. Some atypical SDRs have lost catalytic activity and/or have an unusual NAD(P)-binding motif and missing or unusual active site residues. Reactions catalyzed within the SDR family include isomerization, decarboxylation, epimerization, C=N bond reduction, dehydratase activity, dehalogenation, Enoyl-CoA reduction, and carbonyl-alcohol oxidoreduction.
Pssm-ID: 187586 [Multi-domain] Cd Length: 233 Bit Score: 41.51 E-value: 2.71e-03
Database: CDSEARCH/cdd Low complexity filter: no Composition Based Adjustment: yes E-value threshold: 0.01
References:
Wang J et al. (2023), "The conserved domain database in 2023", Nucleic Acids Res.51(D)384-8.
Lu S et al. (2020), "The conserved domain database in 2020", Nucleic Acids Res.48(D)265-8.
Marchler-Bauer A et al. (2017), "CDD/SPARCLE: functional classification of proteins via subfamily domain architectures.", Nucleic Acids Res.45(D)200-3.
of the residues that compose this conserved feature have been mapped to the query sequence.
Click on the triangle to view details about the feature, including a multiple sequence alignment
of your query sequence and the protein sequences used to curate the domain model,
where hash marks (#) above the aligned sequences show the location of the conserved feature residues.
The thumbnail image, if present, provides an approximate view of the feature's location in 3 dimensions.
Click on the triangle for interactive 3D structure viewing options.
Functional characterization of the conserved domain architecture found on the query.
Click here to see more details.
This image shows a graphical summary of conserved domains identified on the query sequence.
The Show Concise/Full Display button at the top of the page can be used to select the desired level of detail: only top scoring hits
(labeled illustration) or all hits
(labeled illustration).
Domains are color coded according to superfamilies
to which they have been assigned. Hits with scores that pass a domain-specific threshold
(specific hits) are drawn in bright colors.
Others (non-specific hits) and
superfamily placeholders are drawn in pastel colors.
if a domain or superfamily has been annotated with functional sites (conserved features),
they are mapped to the query sequence and indicated through sets of triangles
with the same color and shade of the domain or superfamily that provides the annotation. Mouse over the colored bars or triangles to see descriptions of the domains and features.
click on the bars or triangles to view your query sequence embedded in a multiple sequence alignment of the proteins used to develop the corresponding domain model.
The table lists conserved domains identified on the query sequence. Click on the plus sign (+) on the left to display full descriptions, alignments, and scores.
Click on the domain model's accession number to view the multiple sequence alignment of the proteins used to develop the corresponding domain model.
To view your query sequence embedded in that multiple sequence alignment, click on the colored bars in the Graphical Summary portion of the search results page,
or click on the triangles, if present, that represent functional sites (conserved features)
mapped to the query sequence.
Concise Display shows only the best scoring domain model, in each hit category listed below except non-specific hits, for each region on the query sequence.
(labeled illustration) Standard Display shows only the best scoring domain model from each source, in each hit category listed below for each region on the query sequence.
(labeled illustration) Full Display shows all domain models, in each hit category below, that meet or exceed the RPS-BLAST threshold for statistical significance.
(labeled illustration) Four types of hits can be shown, as available,
for each region on the query sequence:
specific hits meet or exceed a domain-specific e-value threshold
(illustrated example)
and represent a very high confidence that the query sequence belongs to the same protein family as the sequences use to create the domain model
non-specific hits
meet or exceed the RPS-BLAST threshold for statistical significance (default E-value cutoff of 0.01, or an E-value selected by user via the
advanced search options)
the domain superfamily to which the specific and non-specific hits belong
multi-domain models that were computationally detected and are likely to contain multiple single domains
Retrieve proteins that contain one or more of the domains present in the query sequence, using the Conserved Domain Architecture Retrieval Tool
(CDART).
Modify your query to search against a different database and/or use advanced search options