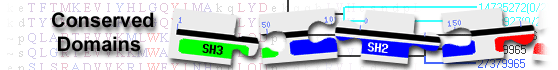


E3 ubiquitin-protein ligase PDZRN3 isoform 1 [Homo sapiens]
Protein Classification

E3 ubiquitin-protein ligase PDZRN3( domain architecture ID 11616446)
E3 ubiquitin-protein ligase PDZRN3 (PDZ domain-containing RING finger protein 3) acts as an E3 ubiquitin-protein ligase that plays an important role in regulating the surface level of MUSK on myotubes
List of domain hits
| Name | Accession | Description | Interval | E-value | |||
| PDZ1-PDZRN4-like | cd06715 | PDZ domain 1 of PDZ domain-containing RING finger protein 4 (PDZRN4), PDZRN3-B, and related ... |
246-337 | 6.28e-59 | |||
PDZ domain 1 of PDZ domain-containing RING finger protein 4 (PDZRN4), PDZRN3-B, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of PDZRN4, PDZRN3-B, and related domains. PDZRN4 (also known as ligand of numb protein X 4, and SEMACAP3-like protein) contains an N-terminal RING domain and two tandem repeat PDZ domains. It is involved in the progression of cancer, including human liver cancer and breast cancer, and may contribute to the tumorigenesis of rectal adenocarcinoma. Danio rerio PDZRN3-B may participate in neurogenesis: the first PDZ domain of Danio rerio Pdzrn3 interacts with Kidins220 (Kinase D-interacting substrate 220 kD, also named Ankyrin Repeat-Rich Membrane Spanning), a crucial mediator of signal transduction in neural tissues. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZRN4-like family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. : Pssm-ID: 467199 [Multi-domain] Cd Length: 92 Bit Score: 196.46 E-value: 6.28e-59
|
|||||||
| PDZ2-PDZRN4-like | cd06716 | PDZ domain 2 of PDZ domain-containing RING finger protein 4 (PDZRN4), PDZRN3-B, and related ... |
420-503 | 7.87e-47 | |||
PDZ domain 2 of PDZ domain-containing RING finger protein 4 (PDZRN4), PDZRN3-B, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of PDZRN4, PDZRN3-B, and related domains. PDZRN4 (also known as ligand of numb protein X 4, and SEMACAP3-like protein) contains an N-terminal RING domain and two tandem repeat PDZ domains. It is involved in the progression of cancer, including human liver cancer and breast cancer, and may contribute to the tumorigenesis of rectal adenocarcinoma. Danio rerio PDZRN3-B may participate in neurogenesis: the first PDZ domain of Danio rerio Pdzrn3 interacts with Kidins220 (Kinase D-interacting substrate 220 kD, also named Ankyrin Repeat-Rich Membrane Spanning), a crucial mediator of signal transduction in neural tissues. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZRN4-like family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. : Pssm-ID: 467200 [Multi-domain] Cd Length: 88 Bit Score: 162.06 E-value: 7.87e-47
|
|||||||
| RING-HC_LNX3 | cd16718 | RING finger, HC subclass, found in ligand of numb protein X 3 (LNX3); LNX3, also known as PDZ ... |
12-58 | 3.15e-31 | |||
RING finger, HC subclass, found in ligand of numb protein X 3 (LNX3); LNX3, also known as PDZ domain-containing RING finger protein 3 (PDZRN3), or Semaphorin cytoplasmic domain-associated protein 3 (SEMACAP3), is an E3 ubiquitin-protein ligase that was first identified as a Semaphorin-binding partner. It is also responsible for the ubiquitination and degradation of Numb, a component of the Notch signaling pathway that functions in the specification of cell fates during development and is known to control cell numbers during neurogenesis in vertebrates. LNX3 acts as a negative regulator of osteoblast differentiation by inhibiting Wnt-beta-catenin signaling. LNX3 also plays an important role in neuromuscular junction formation. It interacts with and ubiquitinates the muscle specific tyrosine kinase (MuSK), thus promoting its endocytosis and negatively regulating the cell surface expression of this key regulator of postsynaptic assembly. LNX3 contains an N-terminal C3HC4-type RING-HC finger, two PDZ domains, and a C-terminal LNX3 homology (LNX3H) domain. : Pssm-ID: 438378 [Multi-domain] Cd Length: 47 Bit Score: 116.24 E-value: 3.15e-31
|
|||||||
| Sina super family | cl47795 | Seven in absentia protein family; The seven in absentia (sina) gene was first identified in ... |
69-132 | 1.13e-06 | |||
Seven in absentia protein family; The seven in absentia (sina) gene was first identified in Drosophila. The Drosophila Sina protein is essential for the determination of the R7 pathway in photoreceptor cell development: the loss of functional Sina results in the transformation of the R7 precursor cell to a non- neuronal cell type. The Sina protein contains an N-terminal RING finger domain pfam00097. Through this domain, Sina binds E2 ubiquitin-conjugating enzymes (UbcD1) Sina also interacts with Tramtrack (TTK88) via PHYL. Tramtrack is a transcriptional repressor that blocks photoreceptor determination, while PHYL down-regulates the activity of TTK88. In turn, the activity of PHYL requires the activation of the Sevenless receptor tyrosine kinase, a process essential for R7 determination. It is thought that thus Sina targets TTK88 for degradation, therefore promoting the R7 pathway. Murine and human homologs of Sina have also been identified. The human homolog Siah-1 also binds E2 enzymes (UbcH5) and through a series of physical interactions, targets beta-catenin for ubiquitin degradation. Siah-1 expression is enhanced by p53, itself promoted by DNA damage. Thus this pathway links DNA damage to beta-catenin degradation. Sina proteins, therefore, physically interact with a variety of proteins. The N-terminal RING finger domain that binds ubiquitin conjugating enzymes is described in pfam00097, and does not form part of the alignment for this family. The remainder C-terminal part is involved in interactions with other proteins, and is included in this alignment. In addition to the Drosophila protein and mammalian homologs, whose similarity was noted previously, this family also includes putative homologs from Caenorhabditis elegans, Arabidopsis thaliana. The actual alignment was detected with superfamily member pfam03145: Pssm-ID: 460824 [Multi-domain] Cd Length: 198 Bit Score: 50.29 E-value: 1.13e-06
|
|||||||
| Name | Accession | Description | Interval | E-value | ||||
| PDZ1-PDZRN4-like | cd06715 | PDZ domain 1 of PDZ domain-containing RING finger protein 4 (PDZRN4), PDZRN3-B, and related ... |
246-337 | 6.28e-59 | ||||
PDZ domain 1 of PDZ domain-containing RING finger protein 4 (PDZRN4), PDZRN3-B, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of PDZRN4, PDZRN3-B, and related domains. PDZRN4 (also known as ligand of numb protein X 4, and SEMACAP3-like protein) contains an N-terminal RING domain and two tandem repeat PDZ domains. It is involved in the progression of cancer, including human liver cancer and breast cancer, and may contribute to the tumorigenesis of rectal adenocarcinoma. Danio rerio PDZRN3-B may participate in neurogenesis: the first PDZ domain of Danio rerio Pdzrn3 interacts with Kidins220 (Kinase D-interacting substrate 220 kD, also named Ankyrin Repeat-Rich Membrane Spanning), a crucial mediator of signal transduction in neural tissues. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZRN4-like family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467199 [Multi-domain] Cd Length: 92 Bit Score: 196.46 E-value: 6.28e-59
|
||||||||
| PDZ2-PDZRN4-like | cd06716 | PDZ domain 2 of PDZ domain-containing RING finger protein 4 (PDZRN4), PDZRN3-B, and related ... |
420-503 | 7.87e-47 | ||||
PDZ domain 2 of PDZ domain-containing RING finger protein 4 (PDZRN4), PDZRN3-B, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of PDZRN4, PDZRN3-B, and related domains. PDZRN4 (also known as ligand of numb protein X 4, and SEMACAP3-like protein) contains an N-terminal RING domain and two tandem repeat PDZ domains. It is involved in the progression of cancer, including human liver cancer and breast cancer, and may contribute to the tumorigenesis of rectal adenocarcinoma. Danio rerio PDZRN3-B may participate in neurogenesis: the first PDZ domain of Danio rerio Pdzrn3 interacts with Kidins220 (Kinase D-interacting substrate 220 kD, also named Ankyrin Repeat-Rich Membrane Spanning), a crucial mediator of signal transduction in neural tissues. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZRN4-like family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467200 [Multi-domain] Cd Length: 88 Bit Score: 162.06 E-value: 7.87e-47
|
||||||||
| RING-HC_LNX3 | cd16718 | RING finger, HC subclass, found in ligand of numb protein X 3 (LNX3); LNX3, also known as PDZ ... |
12-58 | 3.15e-31 | ||||
RING finger, HC subclass, found in ligand of numb protein X 3 (LNX3); LNX3, also known as PDZ domain-containing RING finger protein 3 (PDZRN3), or Semaphorin cytoplasmic domain-associated protein 3 (SEMACAP3), is an E3 ubiquitin-protein ligase that was first identified as a Semaphorin-binding partner. It is also responsible for the ubiquitination and degradation of Numb, a component of the Notch signaling pathway that functions in the specification of cell fates during development and is known to control cell numbers during neurogenesis in vertebrates. LNX3 acts as a negative regulator of osteoblast differentiation by inhibiting Wnt-beta-catenin signaling. LNX3 also plays an important role in neuromuscular junction formation. It interacts with and ubiquitinates the muscle specific tyrosine kinase (MuSK), thus promoting its endocytosis and negatively regulating the cell surface expression of this key regulator of postsynaptic assembly. LNX3 contains an N-terminal C3HC4-type RING-HC finger, two PDZ domains, and a C-terminal LNX3 homology (LNX3H) domain. Pssm-ID: 438378 [Multi-domain] Cd Length: 47 Bit Score: 116.24 E-value: 3.15e-31
|
||||||||
| PDZ | smart00228 | Domain present in PSD-95, Dlg, and ZO-1/2; Also called DHR (Dlg homologous region) or GLGF ... |
246-338 | 5.19e-20 | ||||
Domain present in PSD-95, Dlg, and ZO-1/2; Also called DHR (Dlg homologous region) or GLGF (relatively well conserved tetrapeptide in these domains). Some PDZs have been shown to bind C-terminal polypeptides; others appear to bind internal (non-C-terminal) polypeptides. Different PDZs possess different binding specificities. Pssm-ID: 214570 [Multi-domain] Cd Length: 85 Bit Score: 85.51 E-value: 5.19e-20
|
||||||||
| PDZ | pfam00595 | PDZ domain; PDZ domains are found in diverse signaling proteins. |
249-336 | 4.55e-16 | ||||
PDZ domain; PDZ domains are found in diverse signaling proteins. Pssm-ID: 395476 [Multi-domain] Cd Length: 81 Bit Score: 73.85 E-value: 4.55e-16
|
||||||||
| PDZ | smart00228 | Domain present in PSD-95, Dlg, and ZO-1/2; Also called DHR (Dlg homologous region) or GLGF ... |
431-503 | 3.32e-13 | ||||
Domain present in PSD-95, Dlg, and ZO-1/2; Also called DHR (Dlg homologous region) or GLGF (relatively well conserved tetrapeptide in these domains). Some PDZs have been shown to bind C-terminal polypeptides; others appear to bind internal (non-C-terminal) polypeptides. Different PDZs possess different binding specificities. Pssm-ID: 214570 [Multi-domain] Cd Length: 85 Bit Score: 65.86 E-value: 3.32e-13
|
||||||||
| PDZ | pfam00595 | PDZ domain; PDZ domains are found in diverse signaling proteins. |
425-501 | 9.41e-11 | ||||
PDZ domain; PDZ domains are found in diverse signaling proteins. Pssm-ID: 395476 [Multi-domain] Cd Length: 81 Bit Score: 58.83 E-value: 9.41e-11
|
||||||||
| zf-C3HC4_2 | pfam13923 | Zinc finger, C3HC4 type (RING finger); |
17-53 | 3.72e-09 | ||||
Zinc finger, C3HC4 type (RING finger); Pssm-ID: 404756 [Multi-domain] Cd Length: 40 Bit Score: 53.21 E-value: 3.72e-09
|
||||||||
| CtpA | COG0793 | C-terminal processing protease CtpA/Prc, contains a PDZ domain [Posttranslational modification, ... |
431-503 | 4.26e-07 | ||||
C-terminal processing protease CtpA/Prc, contains a PDZ domain [Posttranslational modification, protein turnover, chaperones]; Pssm-ID: 440556 [Multi-domain] Cd Length: 341 Bit Score: 53.34 E-value: 4.26e-07
|
||||||||
| RING | smart00184 | Ring finger; E3 ubiquitin-protein ligase activity is intrinsic to the RING domain of c-Cbl and ... |
18-53 | 9.24e-07 | ||||
Ring finger; E3 ubiquitin-protein ligase activity is intrinsic to the RING domain of c-Cbl and is likely to be a general function of this domain; Various RING fingers exhibit binding activity towards E2 ubiquitin-conjugating enzymes (Ubc' s) Pssm-ID: 214546 [Multi-domain] Cd Length: 40 Bit Score: 46.35 E-value: 9.24e-07
|
||||||||
| Sina | pfam03145 | Seven in absentia protein family; The seven in absentia (sina) gene was first identified in ... |
69-132 | 1.13e-06 | ||||
Seven in absentia protein family; The seven in absentia (sina) gene was first identified in Drosophila. The Drosophila Sina protein is essential for the determination of the R7 pathway in photoreceptor cell development: the loss of functional Sina results in the transformation of the R7 precursor cell to a non- neuronal cell type. The Sina protein contains an N-terminal RING finger domain pfam00097. Through this domain, Sina binds E2 ubiquitin-conjugating enzymes (UbcD1) Sina also interacts with Tramtrack (TTK88) via PHYL. Tramtrack is a transcriptional repressor that blocks photoreceptor determination, while PHYL down-regulates the activity of TTK88. In turn, the activity of PHYL requires the activation of the Sevenless receptor tyrosine kinase, a process essential for R7 determination. It is thought that thus Sina targets TTK88 for degradation, therefore promoting the R7 pathway. Murine and human homologs of Sina have also been identified. The human homolog Siah-1 also binds E2 enzymes (UbcH5) and through a series of physical interactions, targets beta-catenin for ubiquitin degradation. Siah-1 expression is enhanced by p53, itself promoted by DNA damage. Thus this pathway links DNA damage to beta-catenin degradation. Sina proteins, therefore, physically interact with a variety of proteins. The N-terminal RING finger domain that binds ubiquitin conjugating enzymes is described in pfam00097, and does not form part of the alignment for this family. The remainder C-terminal part is involved in interactions with other proteins, and is included in this alignment. In addition to the Drosophila protein and mammalian homologs, whose similarity was noted previously, this family also includes putative homologs from Caenorhabditis elegans, Arabidopsis thaliana. Pssm-ID: 460824 [Multi-domain] Cd Length: 198 Bit Score: 50.29 E-value: 1.13e-06
|
||||||||
| CtpA | COG0793 | C-terminal processing protease CtpA/Prc, contains a PDZ domain [Posttranslational modification, ... |
242-337 | 1.66e-06 | ||||
C-terminal processing protease CtpA/Prc, contains a PDZ domain [Posttranslational modification, protein turnover, chaperones]; Pssm-ID: 440556 [Multi-domain] Cd Length: 341 Bit Score: 51.41 E-value: 1.66e-06
|
||||||||
| PEX10 | COG5574 | RING-finger-containing E3 ubiquitin ligase [Posttranslational modification, protein turnover, ... |
15-66 | 2.81e-06 | ||||
RING-finger-containing E3 ubiquitin ligase [Posttranslational modification, protein turnover, chaperones]; Pssm-ID: 227861 [Multi-domain] Cd Length: 271 Bit Score: 50.28 E-value: 2.81e-06
|
||||||||
| rad18 | TIGR00599 | DNA repair protein rad18; All proteins in this family for which functions are known are ... |
12-57 | 5.83e-06 | ||||
DNA repair protein rad18; All proteins in this family for which functions are known are involved in nucleotide excision repair.This family is based on the phylogenomic analysis of JA Eisen (1999, Ph.D. Thesis, Stanford University). [DNA metabolism, DNA replication, recombination, and repair] Pssm-ID: 273165 [Multi-domain] Cd Length: 397 Bit Score: 50.00 E-value: 5.83e-06
|
||||||||
| degP_htrA_DO | TIGR02037 | periplasmic serine protease, Do/DeqQ family; This family consists of a set proteins various ... |
277-502 | 6.67e-06 | ||||
periplasmic serine protease, Do/DeqQ family; This family consists of a set proteins various designated DegP, heat shock protein HtrA, and protease DO. The ortholog in Pseudomonas aeruginosa is designated MucD and is found in an operon that controls mucoid phenotype. This family also includes the DegQ (HhoA) paralog in E. coli which can rescue a DegP mutant, but not the smaller DegS paralog, which cannot. Members of this family are located in the periplasm and have separable functions as both protease and chaperone. Members have a trypsin domain and two copies of a PDZ domain. This protein protects bacteria from thermal and other stresses and may be important for the survival of bacterial pathogens.// The chaperone function is dominant at low temperatures, whereas the proteolytic activity is turned on at elevated temperatures. [Protein fate, Protein folding and stabilization, Protein fate, Degradation of proteins, peptides, and glycopeptides] Pssm-ID: 273938 [Multi-domain] Cd Length: 428 Bit Score: 49.91 E-value: 6.67e-06
|
||||||||
| PLN03208 | PLN03208 | E3 ubiquitin-protein ligase RMA2; Provisional |
11-45 | 8.94e-03 | ||||
E3 ubiquitin-protein ligase RMA2; Provisional Pssm-ID: 178747 [Multi-domain] Cd Length: 193 Bit Score: 38.53 E-value: 8.94e-03
|
||||||||
| Name | Accession | Description | Interval | E-value | ||||
| PDZ1-PDZRN4-like | cd06715 | PDZ domain 1 of PDZ domain-containing RING finger protein 4 (PDZRN4), PDZRN3-B, and related ... |
246-337 | 6.28e-59 | ||||
PDZ domain 1 of PDZ domain-containing RING finger protein 4 (PDZRN4), PDZRN3-B, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of PDZRN4, PDZRN3-B, and related domains. PDZRN4 (also known as ligand of numb protein X 4, and SEMACAP3-like protein) contains an N-terminal RING domain and two tandem repeat PDZ domains. It is involved in the progression of cancer, including human liver cancer and breast cancer, and may contribute to the tumorigenesis of rectal adenocarcinoma. Danio rerio PDZRN3-B may participate in neurogenesis: the first PDZ domain of Danio rerio Pdzrn3 interacts with Kidins220 (Kinase D-interacting substrate 220 kD, also named Ankyrin Repeat-Rich Membrane Spanning), a crucial mediator of signal transduction in neural tissues. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZRN4-like family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467199 [Multi-domain] Cd Length: 92 Bit Score: 196.46 E-value: 6.28e-59
|
||||||||
| PDZ2-PDZRN4-like | cd06716 | PDZ domain 2 of PDZ domain-containing RING finger protein 4 (PDZRN4), PDZRN3-B, and related ... |
420-503 | 7.87e-47 | ||||
PDZ domain 2 of PDZ domain-containing RING finger protein 4 (PDZRN4), PDZRN3-B, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of PDZRN4, PDZRN3-B, and related domains. PDZRN4 (also known as ligand of numb protein X 4, and SEMACAP3-like protein) contains an N-terminal RING domain and two tandem repeat PDZ domains. It is involved in the progression of cancer, including human liver cancer and breast cancer, and may contribute to the tumorigenesis of rectal adenocarcinoma. Danio rerio PDZRN3-B may participate in neurogenesis: the first PDZ domain of Danio rerio Pdzrn3 interacts with Kidins220 (Kinase D-interacting substrate 220 kD, also named Ankyrin Repeat-Rich Membrane Spanning), a crucial mediator of signal transduction in neural tissues. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZRN4-like family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467200 [Multi-domain] Cd Length: 88 Bit Score: 162.06 E-value: 7.87e-47
|
||||||||
| RING-HC_LNX3 | cd16718 | RING finger, HC subclass, found in ligand of numb protein X 3 (LNX3); LNX3, also known as PDZ ... |
12-58 | 3.15e-31 | ||||
RING finger, HC subclass, found in ligand of numb protein X 3 (LNX3); LNX3, also known as PDZ domain-containing RING finger protein 3 (PDZRN3), or Semaphorin cytoplasmic domain-associated protein 3 (SEMACAP3), is an E3 ubiquitin-protein ligase that was first identified as a Semaphorin-binding partner. It is also responsible for the ubiquitination and degradation of Numb, a component of the Notch signaling pathway that functions in the specification of cell fates during development and is known to control cell numbers during neurogenesis in vertebrates. LNX3 acts as a negative regulator of osteoblast differentiation by inhibiting Wnt-beta-catenin signaling. LNX3 also plays an important role in neuromuscular junction formation. It interacts with and ubiquitinates the muscle specific tyrosine kinase (MuSK), thus promoting its endocytosis and negatively regulating the cell surface expression of this key regulator of postsynaptic assembly. LNX3 contains an N-terminal C3HC4-type RING-HC finger, two PDZ domains, and a C-terminal LNX3 homology (LNX3H) domain. Pssm-ID: 438378 [Multi-domain] Cd Length: 47 Bit Score: 116.24 E-value: 3.15e-31
|
||||||||
| RING-HC_LNX4 | cd16719 | RING finger, HC subclass, found in ligand of numb protein X 4 (LNX4); LNX4, also known as PDZ ... |
12-65 | 2.15e-24 | ||||
RING finger, HC subclass, found in ligand of numb protein X 4 (LNX4); LNX4, also known as PDZ domain-containing RING finger protein 4 (PDZRN4), or SEMACAP3-like protein (SEMCAP3L), is an E3 ubiquitin-protein ligase responsible for the ubiquitination and degradation of Numb, a component of the Notch signaling pathway that functions in the specification of cell fates during development and is known to control cell numbers during neurogenesis in vertebrates. LNX4 contains an N-terminal C3HC4-type RING-HC finger, two PDZ domains, and a C-terminal LNX3 homology (LNX3H) domain. Pssm-ID: 438379 [Multi-domain] Cd Length: 53 Bit Score: 96.92 E-value: 2.15e-24
|
||||||||
| RING-HC_LNX3-like | cd16512 | RING finger, HC subclass, found in ligand of Numb protein LNX3, LNX4, and similar proteins; ... |
16-58 | 4.61e-24 | ||||
RING finger, HC subclass, found in ligand of Numb protein LNX3, LNX4, and similar proteins; The ligand of Numb protein X (LNX) family, also known as PDZ and RING (PDZRN) family, includes LNX1-5, which can interact with Numb, a key regulator of neurogenesis and neuronal differentiation. LNX5 (also known as PDZK4, or PDZRN4L) shows high sequence homology to LNX3 and LNX4, but it lacks the RING domain. LNX1-4 proteins function as E3 ubiquitin ligases and have a unique domain architecture consisting of an N-terminal RING-HC finger for E3 ubiquitin ligase activity and either two or four PDZ domains necessary for the substrate-binding. This family corresponds to LNX3/LNX4-like proteins, which contains a C3HC4-type RING-HC finger and two PDZ domains. Pssm-ID: 438175 [Multi-domain] Cd Length: 43 Bit Score: 95.56 E-value: 4.61e-24
|
||||||||
| PDZ_canonical | cd00136 | canonical PDZ domain; Canonical PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs ... |
249-336 | 7.35e-23 | ||||
canonical PDZ domain; Canonical PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain. PDZ domains usually bind to short specific peptide sequences located at the C-terminal end of their partner proteins known as PDZ binding motifs. These domains can also interact with internal peptide motifs and certain lipids, and can take part in a head-to-tail oligomerization with other PDZ domains. The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. The canonical PDZ domain contains six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467153 [Multi-domain] Cd Length: 81 Bit Score: 93.38 E-value: 7.35e-23
|
||||||||
| PDZ3_Dlg1-2-4-like | cd06795 | PDZ domain 3 of human discs large homolog 1 (Dlg1), Dlg2, and Dlg4, Drosophila disc large (Dlg) ... |
249-334 | 5.60e-22 | ||||
PDZ domain 3 of human discs large homolog 1 (Dlg1), Dlg2, and Dlg4, Drosophila disc large (Dlg), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 3 of Drosophila Dlg1, human Dlg1, 2, and 4 and related domains. Dlg1 (also known as synapse-associated protein Dlg197; SAP-97), Dlg2 (also known as channel-associated protein of synapse-110; postsynaptic density protein 93, PSD-93), Dlg4 (also known as postsynaptic density protein 95, PSD-95; synapse-associated protein 90, SAP-90) each have 3 PDZ domains and belong to the membrane-associated guanylate kinase family. Dlg1 regulates antigen receptor signaling and cell polarity in lymphocytes, B-cell proliferation and antibody production, and TGFalpha bioavailability; its PDZ3 domain binds pro-TGFalpha, and its PDZ2 domain binds the TACE metalloprotease responsible for cleaving pro-TGFalpha to a soluble form. Dlg2 is involved in N-methyl-D-aspartate (NMDA) receptor signaling, regulating surface expression of NMDA receptors in dorsal horn neurons of the spinal cord; it interacts with NMDA receptor subunits and with Shaker-type K+ channel subunits to cluster into a channel complex. The Dlg4 PDZ1 domain binds NMDA receptors, and its PDZ2 domain binds neuronal nitric oxide synthase (nNOS), forming a complex in neurons. The Drosophila Scribble complex (Scribble, Dlg, and lethal giant larvae) plays a role in apico-basal cell polarity, and in other forms of polarity, including regulation of the actin cytoskeleton, cell signaling and vesicular trafficking, and in tumor development; postsynaptic targeting of Drosophila DLG requires interactions mediated by the first two PDZ domains. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Dlg-like family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467257 [Multi-domain] Cd Length: 91 Bit Score: 91.26 E-value: 5.60e-22
|
||||||||
| PDZ2_Scribble-like | cd06703 | PDZ domain 2 of Drosophila Scribble, human Scribble homolog, and related domains; PDZ (PSD-95 ... |
246-339 | 3.44e-21 | ||||
PDZ domain 2 of Drosophila Scribble, human Scribble homolog, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of Drosophila Scribble (also known as LAP4), human Scribble homolog (also known as hScrib, LAP4, CriB1, ScrB1 and Vartul), and related domains. They belong to the LAP family, which describes proteins that contain either one or four PDZ domains and 16 LRRs (leucine-rich repeats) and function in controlling cell shape, size and subcellular protein localization. In Drosophila, the Scribble complex, comprising Scribble, discs large, and lethal giant larvae, plays a role in apico-basal cell polarity, in other forms of polarity, including regulation of the actin cytoskeleton, cell signaling and vesicular trafficking, and in tumor development. Mammalian Scribble is important in many aspects of cancer development. Scribble and its homologs can be downregulated or overexpressed in cancer; they have a role in cancer beyond their function in loss of cell polarity. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Scribble-like family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467187 [Multi-domain] Cd Length: 92 Bit Score: 88.86 E-value: 3.44e-21
|
||||||||
| PDZ1_Scribble-like | cd06704 | PDZ domain 1 of Drosophila Scribble, human Scribble homolog, and related domains; PDZ (PSD-95 ... |
248-337 | 3.68e-21 | ||||
PDZ domain 1 of Drosophila Scribble, human Scribble homolog, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of Drosophila Scribble (also known as LAP4), human Scribble homolog (also known as hScrib, LAP4, CriB1, ScrB1 and Vartul), and related domains. They belong to the LAP family, which describes proteins that contain either one or four PDZ domains and 16 LRRs (leucine-rich repeats) and function in controlling cell shape, size and subcellular protein localization. In Drosophila, the Scribble complex, comprising Scribble, discs large, and lethal giant larvae, plays a role in apico-basal cell polarity, in other forms of polarity, including regulation of the actin cytoskeleton, cell signaling and vesicular trafficking, and in tumor development. Mammalian Scribble is important in many aspects of cancer development. Scribble and its homologs can be downregulated or overexpressed in cancer; they have a role in cancer beyond their function in loss of cell polarity. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Scribble-like family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467188 [Multi-domain] Cd Length: 87 Bit Score: 88.87 E-value: 3.68e-21
|
||||||||
| PDZ_SYNJ2BP-like | cd06709 | PDZ domain of synaptojanin-2-binding protein (SYNJ2BP), and related domains; PDZ (PSD-95 ... |
248-335 | 5.03e-21 | ||||
PDZ domain of synaptojanin-2-binding protein (SYNJ2BP), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of SYNJ2BP, and related domains. SYNJ2BP (also known as mitochondrial outer membrane protein 25, OMP25) regulates endocytosis of activin type 2 receptor kinases through the Ral/RALBP1-dependent pathway and may be involved in suppression of activin-induced signal transduction. Binding partners of the SYNJ2BP PDZ domain include activin type II receptors (ActR-II), and SYNJ2. SYNJ2BP interacts with the PDZ binding motif of the Notch Delta-like ligand 1 (DLL1) and DLL4, promoting Delta-Notch signaling, and inhibiting sprouting angiogenesis. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This SYNJ2BP-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467193 [Multi-domain] Cd Length: 86 Bit Score: 88.12 E-value: 5.03e-21
|
||||||||
| PDZ3_Scribble-like | cd06702 | PDZ domain 3 of Drosophila Scribble, human Scribble homolog, and related domains; PDZ (PSD-95 ... |
251-337 | 1.67e-20 | ||||
PDZ domain 3 of Drosophila Scribble, human Scribble homolog, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of Drosophila Scribble (also known as LAP4), human Scribble homolog (also known as hScrib, LAP4, CriB1, ScrB1 and Vartul), and related domains. They belong to the LAP family, which describes proteins that contain either one or four PDZ domains and 16 LRRs (leucine-rich repeats) and function in controlling cell shape, size and subcellular protein localization. In Drosophila, the Scribble complex, comprising Scribble, discs large, and lethal giant larvae, plays a role in apico-basal cell polarity, in other forms of polarity, including regulation of the actin cytoskeleton, cell signaling and vesicular trafficking, and in tumor development. Mammalian Scribble is important in many aspects of cancer development. Scribble and its homologs can be downregulated or overexpressed in cancer; they have a role in cancer beyond their function in loss of cell polarity. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Scribble-like family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467186 [Multi-domain] Cd Length: 89 Bit Score: 86.93 E-value: 1.67e-20
|
||||||||
| PDZ | smart00228 | Domain present in PSD-95, Dlg, and ZO-1/2; Also called DHR (Dlg homologous region) or GLGF ... |
246-338 | 5.19e-20 | ||||
Domain present in PSD-95, Dlg, and ZO-1/2; Also called DHR (Dlg homologous region) or GLGF (relatively well conserved tetrapeptide in these domains). Some PDZs have been shown to bind C-terminal polypeptides; others appear to bind internal (non-C-terminal) polypeptides. Different PDZs possess different binding specificities. Pssm-ID: 214570 [Multi-domain] Cd Length: 85 Bit Score: 85.51 E-value: 5.19e-20
|
||||||||
| PDZ7_MUPP1-PD6_PATJ-like | cd06671 | PDZ domain 7 of multi-PDZ-domain protein 1 (MUPP1), PDZ domain 6 of PATJ (protein-associated ... |
252-337 | 1.21e-18 | ||||
PDZ domain 7 of multi-PDZ-domain protein 1 (MUPP1), PDZ domain 6 of PATJ (protein-associated tight junction) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 7 of MUPP1 and PDZ domain 6 of PATJ, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ7 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467159 [Multi-domain] Cd Length: 96 Bit Score: 81.98 E-value: 1.21e-18
|
||||||||
| PDZ_canonical | cd00136 | canonical PDZ domain; Canonical PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs ... |
420-490 | 7.89e-18 | ||||
canonical PDZ domain; Canonical PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain. PDZ domains usually bind to short specific peptide sequences located at the C-terminal end of their partner proteins known as PDZ binding motifs. These domains can also interact with internal peptide motifs and certain lipids, and can take part in a head-to-tail oligomerization with other PDZ domains. The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. The canonical PDZ domain contains six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467153 [Multi-domain] Cd Length: 81 Bit Score: 79.12 E-value: 7.89e-18
|
||||||||
| PDZ2-PTPN13_FRMPD2-like | cd06792 | PDZ domain 2 of tyrosine kinase PTPN13, FERM and PDZ domain-containing protein 2 (FRMPD2), and ... |
249-338 | 1.75e-17 | ||||
PDZ domain 2 of tyrosine kinase PTPN13, FERM and PDZ domain-containing protein 2 (FRMPD2), and similar domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of human PTPN13, and related domains. PTPN13, also known as Fas-associated protein-tyrosine phosphatase 1 (FAP-1), protein-tyrosine phosphatase 1E (PTP-E1), and protein-tyrosine phosphatase (PTPL1), negatively regulates FAS-mediated apoptosis and NGFR-mediated pro-apoptotic signaling, and may also regulate phosphoinositide 3-kinase (PI3K) signaling. It contains 5 PDZ domains; interaction partners of its second PDZ domain (PDZ2) include the Fas receptor (TNFRSF6) and thyroid receptor-interacting protein 6 (TRIP6). The second PDZ (PDZ2) domain, but not PDZ1 or PDZ3, of FRMPD2 binds to GluN2A and GluN2B, two subunits of N-methyl-d-aspartic acid (NMDA) receptors. Other binding partners of the FRMPDZ2 PDZ2 domain include NOD2, and catenin family members, delta catenin (CTNND2), armadillo repeat gene deleted in velo-cardio-facial syndrome (ARVCF) and p0071 (also known as plakophilin 4; PKP4). PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PTPN13-like family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467254 [Multi-domain] Cd Length: 87 Bit Score: 78.41 E-value: 1.75e-17
|
||||||||
| PDZ4_Scribble-like | cd06701 | PDZ domain 4 of Drosophila Scribble, human Scribble homolog, and related domains; PDZ (PSD-95 ... |
237-335 | 3.79e-16 | ||||
PDZ domain 4 of Drosophila Scribble, human Scribble homolog, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 4 of Drosophila Scribble (also known as LAP4), human Scribble homolog (also known as hScrib, LAP4, CriB1, ScrB1 and Vartul), and related domains. They belong to the LAP family, which describes proteins that contain either one or four PDZ domains and 16 LRRs (leucine-rich repeats) and function in controlling cell shape, size and subcellular protein localization. In Drosophila, the Scribble complex, comprising Scribble, discs large, and lethal giant larvae, plays a role in apico-basal cell polarity, in other forms of polarity, including regulation of the actin cytoskeleton, cell signaling and vesicular trafficking, and in tumor development. Mammalian Scribble is important in many aspects of cancer development. Scribble and its homologs can be downregulated or overexpressed in cancer; they have a role in cancer beyond their function in loss of cell polarity. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Scribble-like family PDZ4 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467185 [Multi-domain] Cd Length: 98 Bit Score: 74.95 E-value: 3.79e-16
|
||||||||
| PDZ | pfam00595 | PDZ domain; PDZ domains are found in diverse signaling proteins. |
249-336 | 4.55e-16 | ||||
PDZ domain; PDZ domains are found in diverse signaling proteins. Pssm-ID: 395476 [Multi-domain] Cd Length: 81 Bit Score: 73.85 E-value: 4.55e-16
|
||||||||
| PDZ_TAX1BP3-like | cd10822 | PDZ domain of tax1-binding protein 3 (TAX1BP3), and related domains; PDZ (PSD-95 (Postsynaptic ... |
252-332 | 1.13e-15 | ||||
PDZ domain of tax1-binding protein 3 (TAX1BP3), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of TAX1BP3, and related domains. TAX1BP3 (also known as glutaminase-interacting protein 3, tax interaction protein 1, TIP-1, tax-interacting protein 1) may regulate a number of protein-protein interactions by competing for PDZ domain binding sites. TAX1BP3 binds beta-catenin and may act as an inhibitor of the Wnt signaling pathway. It competes with LIN7A (also known as Lin-7A or LIN-7A) for inward rectifier potassium channel 4 (KCNJ4) binding, and thereby promotes KCNJ4 internalization. It may play a role in the Rho signaling pathway, and in the activation of CDC42 by the viral protein HPV16 E6. Binding partners of the TAX1BP3 PDZ domain include beta-catenin, KCNJ4, glutaminase liver isoform (GLS2), rho guanine nucleotide exchange factor 16 (ARHGEF16), rhotekin, and CDK5 regulatory subunit-associated protein 3 (also known as LAPZ). PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This TAX1BP3-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467265 [Multi-domain] Cd Length: 94 Bit Score: 73.53 E-value: 1.13e-15
|
||||||||
| PDZ3_PDZD2-PDZ1_hPro-IL-16-like | cd06759 | PDZ domain 3 of PDZ domain containing 2 (PDZD2), PDZ domain 1 of human pro-interleukin-16 ... |
246-337 | 9.68e-15 | ||||
PDZ domain 3 of PDZ domain containing 2 (PDZD2), PDZ domain 1 of human pro-interleukin-16 (isoform 1, 1332 AA), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 3 of PDZD2, also known as KIAA0300, PIN-1, activated in prostate cancer (AIPC) and PDZ domain-containing protein 3 (PDZK3). PDZD2 has seven PDZ domains. PDZD2 is expressed at exceptionally high levels in the pancreas and certain cancer tissues, such as prostate cancer. It promotes the proliferation of insulinoma cells and is upregulated during prostate tumorigenesis. In osteosarcoma (OS), the microRNA miR-363 acts as a tumor suppressor by inhibiting PDZD2. This family also includes the first PDZ domain (PDZ1) of human pro-interleukin-16 (isoform 1, also known as nPro-Il-16; 1332 amino-acid protein). Precursor IL-16 is cleaved to produce pro-IL-16 and mature IL-16 (derived from the C-terminal 121 AA). Pro-IL-16 functions as a regulator of T cell growth; mature IL-16 is a CD4 ligand that induces chemotaxis and CD25 expression in CD4+ T cells. IL-16 bioactivity has been closely associated with the progression of several different cancers. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZD2-like family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467240 [Multi-domain] Cd Length: 87 Bit Score: 70.38 E-value: 9.68e-15
|
||||||||
| PDZ2_Dlg1-2-4-like | cd06724 | PDZ domain 2 of human discs large homolog 1 (Dlg1), Dlg2, and Dlg4, Drosophila disc large (Dlg) ... |
250-335 | 1.22e-14 | ||||
PDZ domain 2 of human discs large homolog 1 (Dlg1), Dlg2, and Dlg4, Drosophila disc large (Dlg), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of Drosophila Dlg1, human Dlg1,2, and 4 and related domains. Dlg1 (also known as synapse-associated protein Dlg197 or SAP-97), Dlg2 (also known as channel-associated protein of synapse-110, postsynaptic density protein 93, or PSD-93), Dlg4 (also known as postsynaptic density protein 95, PSD-95, synapse-associated protein 90, or SAP-90) each have 3 PDZ domains and belong to the membrane-associated guanylate kinase family. Dlg1 regulates antigen receptor signaling and cell polarity in lymphocytes, B-cell proliferation and antibody production, and TGFalpha bioavailability; its PDZ3 domain binds pro-TGFalpha, and its PDZ2 domain binds the TACE metalloprotease responsible for cleaving pro-TGFalpha to a soluble form. Dlg2 is involved in N-methyl-D-aspartate (NMDA) receptor signaling. It regulates surface expression of NMDA receptors in dorsal horn neurons of the spinal cord, and it also interacts with NMDA receptor subunits and with Shaker-type K+ channel subunits to cluster into a channel complex. Dlg4 PDZ1 domain binds NMDA receptors, and its PDZ2 domain binds neuronal nitric oxide synthase (nNOS), forming a complex in neurons. The Drosophila Scribble complex (Scribble, Dlg, and lethal giant larvae) plays a role in apico-basal cell polarity, and in other forms of polarity, including regulation of the actin cytoskeleton, cell signaling and vesicular trafficking, and in tumor development. Postsynaptic targeting of Drosophila DLG requires interactions mediated by the first two PDZ domains. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Dlg-like family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467207 [Multi-domain] Cd Length: 85 Bit Score: 69.99 E-value: 1.22e-14
|
||||||||
| PDZ6_PDZD2-PDZ3_hPro-IL-16-like | cd06762 | PDZ domain 6 of PDZ domain containing 2 (PDZD2), PDZ domain 3 of human pro-interleukin-16 ... |
251-338 | 2.75e-14 | ||||
PDZ domain 6 of PDZ domain containing 2 (PDZD2), PDZ domain 3 of human pro-interleukin-16 (isoform 1, 1332 AA), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 6 of PDZD2, also known as KIAA0300, PIN-1, activated in prostate cancer (AIPC) and PDZ domain-containing protein 3 (PDZK3). PDZD2 has seven PDZ domains. PDZD2 is expressed at exceptionally high levels in the pancreas and certain cancer tissues, such as prostate cancer. It promotes the proliferation of insulinoma cells and is upregulated during prostate tumorigenesis. In osteosarcoma (OS), the microRNA miR-363 acts as a tumor suppressor by inhibiting PDZD2. This family also includes the third PDZ domain (PDZ3) of human pro-interleukin-16 (isoform 1, also known as nPro-IL-16). Precursor IL-16 is cleaved to produce pro-IL-16 and C-terminal mature IL-16. Pro-IL-16 functions as a regulator of T cell growth; mature IL-16 is a CD4 ligand that induces chemotaxis and CD25 expression in CD4+ T cells. IL-16 bioactivity has been closely associated with the progression of several different cancers. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZD2-like family PDZ6 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467243 [Multi-domain] Cd Length: 86 Bit Score: 69.21 E-value: 2.75e-14
|
||||||||
| PDZ1_Dlg1-2-4-like | cd06723 | PDZ domain 1 of human discs large homolog 1 (Dlg1), Dlg2, and Dlg4, Drosophila disc large (Dlg) ... |
248-338 | 3.60e-14 | ||||
PDZ domain 1 of human discs large homolog 1 (Dlg1), Dlg2, and Dlg4, Drosophila disc large (Dlg), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of Drosophila Dlg1, human Dlg1,2, and 4 and related domains. Dlg1 (also known as synapse-associated protein Dlg197 or SAP-97), Dlg2 (also known as channel-associated protein of synapse-110, postsynaptic density protein 93, or PSD-93), Dlg4 (also known as postsynaptic density protein 95, PSD-95, synapse-associated protein 90, or SAP-90) each have 3 PDZ domains and belong to the membrane-associated guanylate kinase family. Dlg1 regulates antigen receptor signaling and cell polarity in lymphocytes, B-cell proliferation and antibody production, and TGFalpha bioavailability; its PDZ3 domain binds pro-TGFalpha, and its PDZ2 domain binds the TACE metalloprotease responsible for cleaving pro-TGFalpha to a soluble form. Dlg2 is involved in N-methyl-D-aspartate (NMDA) receptor signaling. It regulates surface expression of NMDA receptors in dorsal horn neurons of the spinal cord, and it also interacts with NMDA receptor subunits and with Shaker-type K+ channel subunits to cluster into a channel complex. Dlg4 PDZ1 domain binds NMDA receptors, and its PDZ2 domain binds neuronal nitric oxide synthase (nNOS), forming a complex in neurons. The Drosophila Scribble complex (Scribble, Dlg, and lethal giant larvae) plays a role in apico-basal cell polarity, and in other forms of polarity, including regulation of the actin cytoskeleton, cell signaling and vesicular trafficking, and in tumor development. Postsynaptic targeting of Drosophila DLG requires interactions mediated by the first two PDZ domains. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Dlg-like family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467206 [Multi-domain] Cd Length: 89 Bit Score: 68.88 E-value: 3.60e-14
|
||||||||
| PDZ13_MUPP1-like | cd06676 | PDZ domain 13 of multi-PDZ-domain protein 1 (MUPP1) and related domains; PDZ (PSD-95 ... |
249-336 | 6.79e-14 | ||||
PDZ domain 13 of multi-PDZ-domain protein 1 (MUPP1) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 13 of MUPP1. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, PDZ9, and PDZ13. This MuPP1-like PDZ13 domain is therefore absent from PATJ. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ13 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467164 [Multi-domain] Cd Length: 83 Bit Score: 67.75 E-value: 6.79e-14
|
||||||||
| PDZ1_PTPN13_FRMPD2-like | cd06694 | PDZ domain 1 of protein tyrosine phosphatase non-receptor type 13 (PTPN13),FERM and PDZ ... |
259-343 | 2.63e-13 | ||||
PDZ domain 1 of protein tyrosine phosphatase non-receptor type 13 (PTPN13),FERM and PDZ domain-containing protein 2 (FRMPD2), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of PTPN13 [also known as Fas-associated protein-tyrosine phosphatase 1 (FAP-1), protein-tyrosine phosphatase 1E (PTP-E1), and protein-tyrosine phosphatase (PTPL1)], FRMPD2 (also known as PDZ domain-containing protein 4; PDZ domain-containing protein 5C), and related domains. PTPN13 regulates negative apoptotic signaling and mediates phosphoinositide 3-kinase (PI3K) signaling. PTPN13 has five PDZ domains. Proteins known to interact with PTPN13 PDZ domains include: PLEKHA1 and PLEKHA2 via PTPN13-PDZ domain 1, Fas receptor and thyroid receptor-interacting protein 6 via PTPN13-PDZ domain 2, nerve growth factor receptor and protein kinase N2 via PTPN13-PDZ domain 3, PDZ and LIM domain 4 (PDLIM4) via PTPN13-PDZ domains 2 and 4, and brain calpain-2 via PTPN13-PDZ domains 3, 4 and 5. Calpain-2-mediated PTPN13 fragments may be involved in abnormal tau aggregation and increased risk for Alzheimer's disease. FRMPD2 is localized in the basolateral membranes of polarized epithelial cells and is associated with tight junction formation and immune response; it contains 3 PDZ domains. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PTPN13 family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467180 [Multi-domain] Cd Length: 92 Bit Score: 66.65 E-value: 2.63e-13
|
||||||||
| PDZ | smart00228 | Domain present in PSD-95, Dlg, and ZO-1/2; Also called DHR (Dlg homologous region) or GLGF ... |
431-503 | 3.32e-13 | ||||
Domain present in PSD-95, Dlg, and ZO-1/2; Also called DHR (Dlg homologous region) or GLGF (relatively well conserved tetrapeptide in these domains). Some PDZs have been shown to bind C-terminal polypeptides; others appear to bind internal (non-C-terminal) polypeptides. Different PDZs possess different binding specificities. Pssm-ID: 214570 [Multi-domain] Cd Length: 85 Bit Score: 65.86 E-value: 3.32e-13
|
||||||||
| PDZ5_DrPTPN13-like | cd23060 | PDZ domain 5 of Danio rerio tyrosine-protein phosphatase non-receptor type 13 (Ptpn13) and ... |
255-328 | 4.90e-13 | ||||
PDZ domain 5 of Danio rerio tyrosine-protein phosphatase non-receptor type 13 (Ptpn13) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 5 of Danio rerio Ptpn13, and related domains. Protein-tyrosine phosphatases (PTPs) dephosphorylate phosphotyrosyl residues in proteins that are phosphorylated by protein tyrosine kinases (PTKs). Danio rerio Ptpn13 is a classical non-receptor-like PTP. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PTPN13-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467273 [Multi-domain] Cd Length: 80 Bit Score: 65.45 E-value: 4.90e-13
|
||||||||
| PDZ1_ZO1-like | cd06727 | PDZ domain 1 of Zonula Occludens-1 (ZO-1), homologs ZO-2 and ZO-3, and related domains; PDZ ... |
249-338 | 5.32e-13 | ||||
PDZ domain 1 of Zonula Occludens-1 (ZO-1), homologs ZO-2 and ZO-3, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of ZO-1, -2, -3 and related domains. Zonula occludens proteins (ZO-1, ZO-2, ZO-3) are multi-PDZ domain proteins involved in the maintenance and biogenesis of multi-protein networks at the cytoplasmic surface of intercellular contacts in epithelial and endothelial cells. They have three N-terminal PDZ domains, PDZ1-3, followed by a Src homology-3 (SH3) domain and a guanylate kinase (GuK)-like domain. Among protein-protein interactions for all ZO proteins is the binding of the first PDZ domain (PDZ1) to the C-termini of claudins, and the homo- and hetero-dimerization of ZO-proteins via their second PDZ domain (PDZ2), which takes place by symmetrical domain swapping of the first two beta-strands of PDZ2. At the cell level, ZO-1 and ZO-2 are involved in polarity maintenance, gene transcription, cell proliferation, and tumor cell metastasis. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This ZO family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467209 [Multi-domain] Cd Length: 87 Bit Score: 65.37 E-value: 5.32e-13
|
||||||||
| PDZ_syntrophin-like | cd06801 | PDZ domain of syntrophins, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), ... |
249-335 | 6.08e-13 | ||||
PDZ domain of syntrophins, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of syntrophins (including alpha-1-syntrophin, beta-1-syntrophin, beta-2-syntrophin, gamma-1-syntrophin, and gamma-2-syntrophin), and related domains. Syntrophins play a role in recruiting various signaling molecules into signaling complexes and help provide appropriate spatiotemporal regulation of signaling pathways. They function in cytoskeletal organization and maintenance; as components of the dystrophin-glycoprotein complex (DGC), they help maintain structural integrity of skeletal muscle fibers. They link voltage-gated sodium channels to the actin cytoskeleton and the extracellular matrix, and control the localization and activity of the actin reorganizing proteins such as PI3K, PI(3,4)P2 and TAPP1. Through association with various cytoskeletal proteins within the cells, they are involved in processes such as regulation of focal adhesions, myogenesis, calcium homeostasis, and cell migration. They also have roles in synapse formation and in the organization of utrophin, acetylcholine receptor, and acetylcholinesterase at the neuromuscular synapse. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This syntrophin-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467262 [Multi-domain] Cd Length: 83 Bit Score: 65.29 E-value: 6.08e-13
|
||||||||
| PDZ_densin_erbin-like | cd06749 | PDZ domain of densin, erbin, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95) ... |
259-337 | 6.29e-13 | ||||
PDZ domain of densin, erbin, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of densin, erbin, and related domains. Densin (also known as leucine-rich repeat-containing protein 7, LRRC7, densin-180, protein LAP1) and erbin (also known as densin-180-like protein, Erbb2-interacting protein, protein LAP2) belong to the LAP (leucine-rich repeat and PDZ domain) family of scaffolding proteins that play roles in the maintenance of cell shape and apical-basal polarity. Densin and erbin are components of the excitatory postsynaptic compartment and are regulators of dendritic morphology and postsynaptic structure. The densin PDZ domain binds CaV1.3 alpha1 subunit, delta-catenin, and MAGUIN-1. Binding partners of the erbin PDZ domain include ErbB receptor tyrosine kinase ErbB2, HTLV-1 Tax1, Cav1.3 Ca2+channels, and constituents of the cadherin:catenin cell adhesion complex, in particular delta-catenin, p0071 and ARVCF. The erbin PDZ domain binds Smad3, a transductor of the TGFbeta pathway, possibly by a novel interface of binding. Erbin and two other LAP proteins (scribble and lano) redundantly regulate epithelial polarity and apical adhesion complex. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This densin and erbin-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged as beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467231 [Multi-domain] Cd Length: 87 Bit Score: 65.43 E-value: 6.29e-13
|
||||||||
| PDZ1_GgSTXBP4-like | cd06692 | PDZ1 domain of Gallus gallus uncharacterized syntaxin-binding protein 4 (STXBP4) isoform X1, ... |
426-504 | 6.96e-13 | ||||
PDZ1 domain of Gallus gallus uncharacterized syntaxin-binding protein 4 (STXBP4) isoform X1, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of Gallus gallus uncharacterized syntaxin-binding protein 4 (STXBP4) isoform X1, and related domains. Gallus gallus STXBP4 isoform X1 contains 2 PDZ domains (PDZ1 and PDZ2). PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This STXBP4-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467179 [Multi-domain] Cd Length: 88 Bit Score: 65.32 E-value: 6.96e-13
|
||||||||
| PDZ1_INAD-like | cd23063 | PDZ domain 1 of inactivation-no-after-potential D (INAD), and related domains; PDZ (PSD-95 ... |
258-335 | 1.78e-12 | ||||
PDZ domain 1 of inactivation-no-after-potential D (INAD), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of INAD, and related domains. INAD assembles key enzymes of the Drosophila compound eye photo-transduction pathway into a supramolecular complex, supporting efficient and fast light signaling. It contains 5 PDZ domains arranged in tandem (PDZ1-PDZ5) which independently bind various proteins. INAD PDZ2 binds eye-specific protein kinase C, INAD PDZ3 binds transient receptor potential (TRP) channel, and INAD PDZ4,5 tandem binds NORPA (phospholipase Cbeta, PLCbeta). Mutations of the inaD gene that lead to disruption of each of these interactions impair fly photo signal transduction. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This INAD-like family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467276 [Multi-domain] Cd Length: 87 Bit Score: 64.07 E-value: 1.78e-12
|
||||||||
| PDZ2_Scribble-like | cd06703 | PDZ domain 2 of Drosophila Scribble, human Scribble homolog, and related domains; PDZ (PSD-95 ... |
437-502 | 3.36e-12 | ||||
PDZ domain 2 of Drosophila Scribble, human Scribble homolog, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of Drosophila Scribble (also known as LAP4), human Scribble homolog (also known as hScrib, LAP4, CriB1, ScrB1 and Vartul), and related domains. They belong to the LAP family, which describes proteins that contain either one or four PDZ domains and 16 LRRs (leucine-rich repeats) and function in controlling cell shape, size and subcellular protein localization. In Drosophila, the Scribble complex, comprising Scribble, discs large, and lethal giant larvae, plays a role in apico-basal cell polarity, in other forms of polarity, including regulation of the actin cytoskeleton, cell signaling and vesicular trafficking, and in tumor development. Mammalian Scribble is important in many aspects of cancer development. Scribble and its homologs can be downregulated or overexpressed in cancer; they have a role in cancer beyond their function in loss of cell polarity. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Scribble-like family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467187 [Multi-domain] Cd Length: 92 Bit Score: 63.43 E-value: 3.36e-12
|
||||||||
| PDZ5_MAGI-1_3-like | cd06735 | PDZ domain 5 of membrane-associated guanylate kinase inverted 1 (MAGI-1), MAGI-2, and MAGI-3, ... |
249-338 | 3.67e-12 | ||||
PDZ domain 5 of membrane-associated guanylate kinase inverted 1 (MAGI-1), MAGI-2, and MAGI-3, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 5 of MAGI1, 2, 3 (MAGI is also known as Membrane-associated guanylate kinase, WW and PDZ domain-containing protein) and related domains. MAGI proteins have been implicated in the control of cell migration and invasion through altering the activity of phosphatase and tensin homolog (PTEN) and modulating Akt signaling. Four MAGI proteins have been identified (MAGI1-3 and MAGIX). MAGI1-3 have 6 PDZ domains and bind to the C-terminus of PTEN via their PDZ2 domain. MAGIX has a single PDZ domain that is related to MAGI1-3 PDZ domain 5, and belongs to this MAGI1,2,3-like family. Other binding partners for MAGI1 include JAM4, C-terminal tail of high risk HPV-18 E6, megalin, TRAF6, Kir4.1 (basolateral K+ channel subunit), and cadherin 23; for MAGI2, include DASM1, dendrin, axin, beta- and delta-catenin, neuroligin, hyperpolarization-activated cation channels, beta1-adrenergic receptors, NMDA receptor, and TARPs; and for MAGI3 includes LPA2. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MAGI family PDZ5 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged as beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467217 [Multi-domain] Cd Length: 84 Bit Score: 62.98 E-value: 3.67e-12
|
||||||||
| PDZ3_MUPP1-like | cd06791 | PDZ domain 3 of multi-PDZ-domain protein 1 (MUPP1) and PATJ (protein-associated tight junction) ... |
252-337 | 6.62e-12 | ||||
PDZ domain 3 of multi-PDZ-domain protein 1 (MUPP1) and PATJ (protein-associated tight junction) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 3 of MUPP1 and PATJ, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467253 [Multi-domain] Cd Length: 89 Bit Score: 62.25 E-value: 6.62e-12
|
||||||||
| PDZ1_GgSTXBP4-like | cd06692 | PDZ1 domain of Gallus gallus uncharacterized syntaxin-binding protein 4 (STXBP4) isoform X1, ... |
259-327 | 7.22e-12 | ||||
PDZ1 domain of Gallus gallus uncharacterized syntaxin-binding protein 4 (STXBP4) isoform X1, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of Gallus gallus uncharacterized syntaxin-binding protein 4 (STXBP4) isoform X1, and related domains. Gallus gallus STXBP4 isoform X1 contains 2 PDZ domains (PDZ1 and PDZ2). PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This STXBP4-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467179 [Multi-domain] Cd Length: 88 Bit Score: 62.24 E-value: 7.22e-12
|
||||||||
| PDZ10_MUPP1-PDZ8_PATJ-like | cd06673 | PDZ domain 10 of multi-PDZ-domain protein 1 (MUPP1), domain 8 of PATJ (protein-associated ... |
242-340 | 7.39e-12 | ||||
PDZ domain 10 of multi-PDZ-domain protein 1 (MUPP1), domain 8 of PATJ (protein-associated tight junction) and similar domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 10 of MUPP1, PDZ domain 8 of PATJ, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ10 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467161 [Multi-domain] Cd Length: 86 Bit Score: 62.31 E-value: 7.39e-12
|
||||||||
| PDZ_Lin-7-like | cd06796 | PDZ domain of protein Lin-7 and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), ... |
259-340 | 1.10e-11 | ||||
PDZ domain of protein Lin-7 and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of Lin-7 (also known as LIN-7 or LIN7), and related domains. Lin-7 targets and organize protein complexes to epithelial and synaptic plasma membranes. There are three mammalian Lin-7 homologs: Lin-7A (protein lin-7 homolog A, also known as mammalian lin-seven protein 1 (MALS-1), vertebrate lin-7 homolog 1 (Veli-1), tax interaction protein 33); Lin-7B (also known as MALS-2, Veli-2); and Lin-7C (also known as MALS-3, Veli-3). Lin-7 is involved in localization of the Let-23 growth factor receptor to the basolateral membrane of epithelial cells, in tight junction localization of insulin receptor substrate p53 (IRSp53), in retaining gamma-aminobutyric (GABA) transporter (BGT-1) at the basolateral surface of epithelial cells, and in regulating recruitment of neurotransmitter receptors to the postsynaptic density (PSD). The Lin7 PDZ domain binds Let-23, BGT and beta-catenin, and NMDA (N-methyl-D-aspartate) receptor NR2B. Lin-7 also binds to the PDZ binding motif located in the C-terminal tail of Rhotekin, an effector protein for small GTPase Rho. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Lin-7-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467258 [Multi-domain] Cd Length: 86 Bit Score: 61.68 E-value: 1.10e-11
|
||||||||
| PDZ3_Par3-like | cd23059 | PDZ domain 3 of partitioning defective 3 (Par3), and related domains; PDZ (PSD-95 ... |
243-338 | 1.23e-11 | ||||
PDZ domain 3 of partitioning defective 3 (Par3), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 3 of Par3 (or PAR3 or Par-3, also known as Atypical PKC isotype-specific-interacting protein, ASIP, Drosophila Bazooka) and related domains. Par3 is a scaffold protein involved in organizing cell polarity across animals. Par3 binds numerous molecules both for its recruitment to one pole of the cell and for downstream contributions to polarized cell function. It regulates cell polarity by targeting the Par complex proteins Par6 and atypical protein kinase C (aPKC) to specific cortical sites. Physical interactions between Par-3 and the Par complex include Par3 PDZ domain 1 binding to the Par6 PDZ domain, Par3 PDZ domain 1 and PDZ domain 3 binding the Par6's PDZ-binding motif, and an interaction with an undefined region of aPKC that requires both Par3 PDZ2 and PDZ3. The PDZ domains of Par3 have also been implicated as potential phosphoinositide signaling integrators, since its second PDZ domain binds to phosphoinositides, and the third PDZ interacts with phosphoinositide phosphatase PTEN. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Par3 family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467272 [Multi-domain] Cd Length: 103 Bit Score: 62.30 E-value: 1.23e-11
|
||||||||
| PDZ4_PDZD2-PDZ2_hPro-IL-16-like | cd06760 | PDZ domain 4 of PDZ domain containing 2 (PDZD2), PDZ domain 2 of human pro-interleukin-16 ... |
431-502 | 1.84e-11 | ||||
PDZ domain 4 of PDZ domain containing 2 (PDZD2), PDZ domain 2 of human pro-interleukin-16 (isoform 1, 1332 AA), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 4 of PDZD2, also known as KIAA0300, PIN-1, activated in prostate cancer (AIPC) and PDZ domain-containing protein 3 (PDZK3). PDZD2 has seven PDZ domains. PDZD2 is expressed at exceptionally high levels in the pancreas and certain cancer tissues, such as prostate cancer. It promotes the proliferation of insulinoma cells and is upregulated during prostate tumorigenesis. In osteosarcoma (OS), the microRNA miR-363 acts as a tumor suppressor by inhibiting PDZD2. This family also includes the second PDZ domain (PDZ2) of human pro-interleukin-16 (isoform 1, also known as nPro-Il-16; 1332 amino-acid protein). Precursor IL-16 is cleaved to produce pro-IL-16 and mature IL-16 (derived from the C-terminal 121 AA). Pro-IL-16 functions as a regulator of T cell growth; mature IL-16 is a CD4 ligand that induces chemotaxis and CD25 expression in CD4+ T cells. IL-16 bioactivity has been closely associated with the progression of several different cancers PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZD2-like family PDZ4 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467241 [Multi-domain] Cd Length: 90 Bit Score: 61.13 E-value: 1.84e-11
|
||||||||
| PDZ2_Par3-like | cd23058 | PDZ domain 2 of partitioning defective 3 (Par3), and related domains; PDZ (PSD-95 ... |
431-502 | 2.79e-11 | ||||
PDZ domain 2 of partitioning defective 3 (Par3), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of Par3 (or PAR3 or Par-3, also known as Atypical PKC isotype-specific-interacting protein, ASIP, Drosophila Bazooka) and related domains. Par3 is a scaffold protein involved in organizing cell polarity across animals. Par3 binds numerous molecules both for its recruitment to one pole of the cell and for downstream contributions to polarized cell function. It regulates cell polarity by targeting the Par complex proteins Par6 and atypical protein kinase C (aPKC) to specific cortical sites. Physical interactions between Par3 and the Par complex include Par3 PDZ domain 1 binding to the Par6 PDZ domain, Par3 PDZ domain 1 and PDZ domain 3 binding the Par6's PDZ-binding motif, and an interaction with an undefined region of aPKC that requires both Par3 PDZ2 and PDZ3. The PDZ domains of Par3 have also been implicated as potential phosphoinositide signaling integrators, since its second PDZ domain binds to phosphoinositides, and the third PDZ interacts with phosphoinositide phosphatase PTEN. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Par3 family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467271 [Multi-domain] Cd Length: 93 Bit Score: 60.73 E-value: 2.79e-11
|
||||||||
| PDZ_Dishevelled-like | cd06717 | PDZ domain of segment polarity protein dishevelled homolog DVL1, DVL2, DVL3, and related ... |
425-488 | 4.84e-11 | ||||
PDZ domain of segment polarity protein dishevelled homolog DVL1, DVL2, DVL3, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of DVL1-3, and related domains. The dishevelleds (DVL1, 2 and 3 in humans) act downstream of Frizzled (FZD) receptors in both the canonical and non-canonical WNT signaling pathway; they bind the cytoplasmic C-terminus of frizzled family members and transduce the Wnt signal to down-stream effectors. They bind to several proteins known to modulate Wnt signaling. Binding partners of the DVL1 PDZ domain include nucleoredoxin (NXN), Van Gogh-like (VANGL1), Wnt receptor RYK, Dapper 1 (DACT1), Frizzled7 (FZD7), transmembrane protein 88 (TMEM88), Daple (dishevelled-associating protein with a high frequency of leucines), also known as Ccdc88c), and cysteine-rich protein Idax. The DVL2 PDZ domain has been shown to bind the nuclear export signal sequence of the DVL2 protein. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This DVL-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467201 [Multi-domain] Cd Length: 87 Bit Score: 60.07 E-value: 4.84e-11
|
||||||||
| PDZ_PDZD11-like | cd06752 | PDZ domain of PDZ domain-containing protein 11, and related domains; PDZ (PSD-95 (Postsynaptic ... |
249-329 | 6.64e-11 | ||||
PDZ domain of PDZ domain-containing protein 11, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of PDZD11, and related domains. PDZD11 (also known as ATPase-interacting PDZ protein, plasma membrane calcium ATPase-interacting single-PDZ protein, PMCA-interacting single-PDZ protein, PISP) is involved in the dynamic assembly of apical junctions (AJs). It is recruited by PLEKHA7 to AJs to promote the efficient junctional recruitment and stabilization of nectins, and the efficient early phases of assembly of AJs in epithelial cells. The PDZD11 PDZ domain binds nectin-1 and nectin-3. PDZD11 also binds to a PDZ binding motif located in the C-terminal tail of the human sodium-dependent multivitamin transporter, to the cytoplasmic tail of the Menkes copper ATPase ATP7A, and to the cytoplasmic tail of all plasma membrane Ca2+-ATPase b-splice variants. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZD11-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467234 [Multi-domain] Cd Length: 83 Bit Score: 59.25 E-value: 6.64e-11
|
||||||||
| PDZ | pfam00595 | PDZ domain; PDZ domains are found in diverse signaling proteins. |
425-501 | 9.41e-11 | ||||
PDZ domain; PDZ domains are found in diverse signaling proteins. Pssm-ID: 395476 [Multi-domain] Cd Length: 81 Bit Score: 58.83 E-value: 9.41e-11
|
||||||||
| PDZ2_GRIP1-2-like | cd06681 | PDZ domain 2 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related ... |
246-325 | 1.21e-10 | ||||
PDZ domain 2 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR) binding proteins GRIP1 (ABP/GRIP2) and GRIP2, and related domains. GRIP1 and GRIP2 each have 7 PDZ domains. The interaction of GRIP1 and GRIP2 with GluA2/3 (AMPAR subunit) regulates AMPAR trafficking and synaptic targeting. GRIP1 has an essential role in regulating AMPAR trafficking during synaptic plasticity and learning and memory. GRIP1 and GRIP2 interact with a variety of other proteins associated with protein trafficking and internalization, for example GRIP1 also interacts with KIF5 (also known as kinesin 1), EphB receptors, scaffold protein liprin-alpha, and the rasGEF GRASP-1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This GRIP family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467169 [Multi-domain] Cd Length: 89 Bit Score: 58.78 E-value: 1.21e-10
|
||||||||
| PDZ12_MUPP1-like | cd06675 | PDZ domain 12 of multi-PDZ-domain protein 1 (MUPP1), PDZ domain 10 of protein-associated tight ... |
249-336 | 1.59e-10 | ||||
PDZ domain 12 of multi-PDZ-domain protein 1 (MUPP1), PDZ domain 10 of protein-associated tight junction (PATJ, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 12 of MUPP1, PDZ domain 10 of PATJ, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like PDZ12 family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F Pssm-ID: 467163 [Multi-domain] Cd Length: 86 Bit Score: 58.53 E-value: 1.59e-10
|
||||||||
| PDZ_6 | pfam17820 | PDZ domain; This entry represents the PDZ domain from a wide variety of proteins. |
448-502 | 1.64e-10 | ||||
PDZ domain; This entry represents the PDZ domain from a wide variety of proteins. Pssm-ID: 436067 [Multi-domain] Cd Length: 54 Bit Score: 57.15 E-value: 1.64e-10
|
||||||||
| PDZ4_LNX1_2-like | cd06680 | PDZ domain 4 of human Ligand of Numb protein X 1 (LNX1) and LNX2, and related domains; PDZ ... |
249-325 | 1.82e-10 | ||||
PDZ domain 4 of human Ligand of Numb protein X 1 (LNX1) and LNX2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 4 of LNX1 (also known as PDZ domain-containing RING finger protein 2, PDZRN2)and LNX2 (also known as PDZ domain-containing RING finger protein 1, PDZRN1), and related domains. LNX1 and LNX2 are Ring (Really Interesting New Gene) finger and PDZ domain-containing E3 ubiquitin ligases that bind to the cell fate determinant protein NUMB and mediate its ubiquitination. LNX1 can ubiquitinate a number of other ligands including PPFIA1, KLHL11, KIF7 and ERC2. LNX1 and LNX2 each have four PDZ domains. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This LNX family PDZ4 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467168 [Multi-domain] Cd Length: 89 Bit Score: 58.51 E-value: 1.82e-10
|
||||||||
| PDZ2_PDZD2-like | cd06758 | PDZ domain 2 of PDZ domain containing 2 (PDZD2), and related domains; PDZ (PSD-95 ... |
252-338 | 1.84e-10 | ||||
PDZ domain 2 of PDZ domain containing 2 (PDZD2), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of PDZD2, also known as KIAA0300, PIN-1, activated in prostate cancer (AIPC) and PDZ domain-containing protein 3 (PDZK3). PDZD2 has seven PDZ domains, and is expressed at exceptionally high levels in the pancreas and certain cancer tissues such as prostate cancer. It promotes the proliferation of insulinoma cells and is upregulated during prostate tumorigenesis. In osteosarcoma (OS), the microRNA miR-363 acts as a tumor suppressor by inhibiting PDZD2. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZD2-like family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467239 [Multi-domain] Cd Length: 88 Bit Score: 58.13 E-value: 1.84e-10
|
||||||||
| PDZ1_FL-whirlin | cd06740 | PDZ domain 1 of the full-length isoform of whirlin and related domains; PDZ (PSD-95 ... |
245-328 | 3.41e-10 | ||||
PDZ domain 1 of the full-length isoform of whirlin and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of the full-length isoform of whirlin and related domains. Whirlin is an essential protein for developmental pathways in photoreceptor cells of the retina and hair cells of the inner ear. The full-length whirlin isoform has two harmonin N-like domains, three PDZ domains, a proline-rich region, and a PDZ-binding motif. Whirlin isoforms may form different complexes at the periciliary membrane complex (PMC) in photoreceptors, and the stereociliary tip and base in inner ear hair cells. It interacts with ADGRV1 and usherin at the PMC; with SANS and RpgrORF15 at the connecting cilium in photoreceptors; with EPS8, MYO15A, p55, and CASK proteins at the stereociliary tip of inner ear hair cells; and with ADGRV1, usherin, and PDZD7 at the stereociliary base in inner ear hair cells. Mutations in the gene encoding whirlin (WHRN; also known as USH2D and DFNB31), have been found to cause either USH2 subtype (USH2D) or autosomal recessive non-syndromic deafness type 31 (DFNB31). Whirlin is the key protein in the USH2 complex (whirlin, usherin and GPR98) which recruits other USH2 causative proteins at the periciliary membrane in photoreceptors and the ankle link of the stereocilia in hair cells. Whirlin's interaction with espin, another stereociliary protein, may be important for the architecture of the USH2 complex. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This whirlin family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467222 [Multi-domain] Cd Length: 82 Bit Score: 57.37 E-value: 3.41e-10
|
||||||||
| RING-HC_ORTHRUS_rpt1 | cd23138 | first RING finger, HC subclass, found in Arabidopsis thaliana ORTHRUS and similar proteins; ... |
15-57 | 3.60e-10 | ||||
first RING finger, HC subclass, found in Arabidopsis thaliana ORTHRUS and similar proteins; This subfamily includes Arabidopsis thaliana ORTHRUS 1-5. They are E3 ubiquitin-protein ligases that may participate in CpG methylation-dependent transcriptional regulation and/or epigenetic transcriptional silencing. ORTHRUS 1 mediates ubiquitination with the E2 ubiquitin-conjugating enzymes UBC11, UBC8 and UBC8 homologs (e.g. UBC10, UBC11, UBC28 and UBC29) but not with UBC27, UBC30, UBC32, UBC34 and UBC36. ORTHRUS 2 and 5 mediate ubiquitination with the E2 ubiquitin-conjugating enzyme UBC11. ORTHRUS 1 and 2 promote methylation-mediated gene silencing leading, for example, to early flowering. They can bind to CpG, CpNpG, and CpNpN DNA motifs, with a strong preference for methylated forms, and with highest affinity for CpG substrates. Members of this subfamily contain two typical C3HC4-type RING-HC fingers. This model corresponds to the first one. Pssm-ID: 438500 [Multi-domain] Cd Length: 48 Bit Score: 56.30 E-value: 3.60e-10
|
||||||||
| PDZ2_harmonin | cd06738 | PDZ domain 2 of harmonin isoforms a, b, and c, and related domains; PDZ (PSD-95 (Postsynaptic ... |
256-329 | 4.27e-10 | ||||
PDZ domain 2 of harmonin isoforms a, b, and c, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of harmonin isoforms a, b, and c, and related domains. Harmonin (also known as Usher Type 1C, PDZ-73 and AIE-75) is a key organizer of the Usher (USH) protein interactome. USH syndrome is the leading cause of hereditary sensory deaf-blindness in humans; three clinically distinct types of USH have been identified, type 1 to 3. The gene encoding harmonin (USH1C) is the causative gene for the USH type 1C phenotype. There are at least 10 alternatively spliced isoforms of harmonin, which are divided into three subclasses (a, b, and c). All isoforms contain the first two PDZ domains and the first coiled-coil domain. The a and b isoforms all have a third PDZ domain. The different PDZ domains are responsible for interactions with all known Usher syndrome type 1 proteins, and most Usher syndrome type 2 proteins. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This harmonin family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467220 [Multi-domain] Cd Length: 82 Bit Score: 56.94 E-value: 4.27e-10
|
||||||||
| PDZ2_FL-whirlin | cd06741 | PDZ domain 2 of the full-length isoform of whirlin and related domains; PDZ (PSD-95 ... |
255-329 | 4.56e-10 | ||||
PDZ domain 2 of the full-length isoform of whirlin and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of the full-length isoform of whirlin and related domains. Whirlin is an essential protein for developmental pathways in photoreceptor cells of the retina and hair cells of the inner ear. The full-length whirlin isoform has two harmonin N-like domains, three PDZ domains, a proline-rich region, and a PDZ-binding motif. Whirlin isoforms may form different complexes at the periciliary membrane complex (PMC) in photoreceptors, and the stereociliary tip and base in inner ear hair cells. It interacts with ADGRV1 and usherin at the PMC; with SANS and RpgrORF15 at the connecting cilium in photoreceptors; with EPS8, MYO15A, p55, and CASK proteins at the stereociliary tip of inner ear hair cells; and with ADGRV1, usherin, and PDZD7 at the stereociliary base in inner ear hair cells. Mutations in the gene encoding whirlin (WHRN; also known as USH2D and DFNB31), have been found to cause either USH2 subtype (USH2D) or autosomal recessive non-syndromic deafness type 31 (DFNB31). Whirlin is the key protein in the USH2 complex (whirlin, usherin and GPR98) which recruits other USH2 causative proteins at the periciliary membrane in photoreceptors and the ankle link of the stereocilia in hair cells. Whirlin's interaction with espin, another stereociliary protein, may be important for the architecture of the USH2 complex. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This whirlin family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467223 [Multi-domain] Cd Length: 84 Bit Score: 56.89 E-value: 4.56e-10
|
||||||||
| PDZ1_PTPN13-like | cd23072 | PDZ domain 1 of protein tyrosine phosphatase non-receptor type 13 (PTPN13), and related ... |
259-335 | 4.64e-10 | ||||
PDZ domain 1 of protein tyrosine phosphatase non-receptor type 13 (PTPN13), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of PTPN13 [also known as Fas-associated protein-tyrosine phosphatase 1 (FAP-1), protein-tyrosine phosphatase 1E (PTP-E1), and protein-tyrosine phosphatase (PTPL1)], and related domains. PTPN13 regulates negative apoptotic signaling and mediates phosphoinositide 3-kinase (PI3K) signaling. PTPN13 has five PDZ domains. Proteins known to interact with PTPN13 PDZ domains include: PLEKHA1 and PLEKHA2 via PTPN13-PDZ domain 1, Fas receptor and thyroid receptor-interacting protein 6 via PTPN13-PDZ domain 2, nerve growth factor receptor and protein kinase N2 via PTPN13-PDZ domain 3, PDZ and LIM domain 4 (PDLIM4) via PTPN13-PDZ domains 2 and 4, and brain calpain-2 via PTPN13-PDZ domains 3, 4 and 5. Calpain-2-mediated PTPN13 fragments may be involved in abnormal tau aggregation and increased risk for Alzheimer's disease. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PTPN13 family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467285 [Multi-domain] Cd Length: 92 Bit Score: 57.50 E-value: 4.64e-10
|
||||||||
| PDZ_Par6-like | cd06718 | PDZ domain of partitioning defective 6 (Par6), Drosophila Rho GTPase-activating protein 100F ... |
254-321 | 5.91e-10 | ||||
PDZ domain of partitioning defective 6 (Par6), Drosophila Rho GTPase-activating protein 100F (RhoGAP100F), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of Par6 (also known as PAR6 or Par-6), RhoGAP100F, and related domains. Par6 is part of a conserved machinery that directs metazoan cell polarity, a process necessary for the function of diverse cell types. Par6 forms a cell polarity-regulatory complex with atypical protein kinase C (aPKC) and Par3. Par6 can also directly associate with PALS1 (proteins associated with Lin7, also known as Stardust) providing a link between the Par3/aPKC/Par6 complex and the PALS1-PATJ (protein-associated TJ) complex. Binding partners of the Par6-PDZ domain include Par3, PALS1/Stardust; leucine-rich repeat-containing protein netrin-G ligand-2 (NGL-2), human crumbs (CRB3) involve in the morphogenesis of the tight junctions in mammalian epithelial cells, and PAR-6 co-operates with the Par6 semi-CRIB domain to bind CDC42. CDC42 regulates the Par6 PDZ domain through an allosteric CRIB-PDZ transition. Drosophila RhoGAP100F, also known as synapse defective protein 1 homolog (syd-1 homolog), is a GTPase activator for the Rho-type GTPases by converting them to an inactive GDP-bound form. The RhoGAP100F-PDZ domain binds the neurexin C terminus to control synapse formation at the Drosophila neuromuscular junction. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Par6-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467202 [Multi-domain] Cd Length: 84 Bit Score: 56.81 E-value: 5.91e-10
|
||||||||
| PDZ3_ZO1-like_domain | cd06729 | PDZ domain 3 of Zonula Occludens-1 (ZO-1), homologs ZO-2 and ZO-3, and related domains; PDZ ... |
444-488 | 7.44e-10 | ||||
PDZ domain 3 of Zonula Occludens-1 (ZO-1), homologs ZO-2 and ZO-3, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 3 of ZO-1, -2, -3 and related domains. Zonula occludens proteins (ZO-1, ZO-2, ZO-3) are multi-PDZ domain proteins involved in the maintenance and biogenesis of multi-protein networks at the cytoplasmic surface of intercellular contacts in epithelial and endothelial cells. They have three N-terminal PDZ domains, PDZ1-3, followed by a Src homology-3 (SH3) domain and a guanylate kinase (GuK)-like domain. Among protein-protein interactions for all ZO proteins is the binding of the first PDZ domain (PDZ1) to the C-termini of claudins , and the homo- and hetero-dimerization of ZO-proteins via their second PDZ domain (PDZ2), which takes place by symmetrical domain swapping of the first two beta-strands of PDZ2. At the cell level, ZO-1 and ZO-2 are involved in polarity maintenance, gene transcription, cell proliferation, and tumor cell metastasis. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This ZO family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467211 [Multi-domain] Cd Length: 82 Bit Score: 56.42 E-value: 7.44e-10
|
||||||||
| PDZ2-PTPN13_FRMPD2-like | cd06792 | PDZ domain 2 of tyrosine kinase PTPN13, FERM and PDZ domain-containing protein 2 (FRMPD2), and ... |
446-488 | 1.19e-09 | ||||
PDZ domain 2 of tyrosine kinase PTPN13, FERM and PDZ domain-containing protein 2 (FRMPD2), and similar domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of human PTPN13, and related domains. PTPN13, also known as Fas-associated protein-tyrosine phosphatase 1 (FAP-1), protein-tyrosine phosphatase 1E (PTP-E1), and protein-tyrosine phosphatase (PTPL1), negatively regulates FAS-mediated apoptosis and NGFR-mediated pro-apoptotic signaling, and may also regulate phosphoinositide 3-kinase (PI3K) signaling. It contains 5 PDZ domains; interaction partners of its second PDZ domain (PDZ2) include the Fas receptor (TNFRSF6) and thyroid receptor-interacting protein 6 (TRIP6). The second PDZ (PDZ2) domain, but not PDZ1 or PDZ3, of FRMPD2 binds to GluN2A and GluN2B, two subunits of N-methyl-d-aspartic acid (NMDA) receptors. Other binding partners of the FRMPDZ2 PDZ2 domain include NOD2, and catenin family members, delta catenin (CTNND2), armadillo repeat gene deleted in velo-cardio-facial syndrome (ARVCF) and p0071 (also known as plakophilin 4; PKP4). PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PTPN13-like family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467254 [Multi-domain] Cd Length: 87 Bit Score: 56.07 E-value: 1.19e-09
|
||||||||
| PDZ9_MUPP1-like | cd10817 | PDZ domain 9 of multi-PDZ-domain protein 1 (MUPP1) and related domains; PDZ (PSD-95 ... |
249-328 | 1.64e-09 | ||||
PDZ domain 9 of multi-PDZ-domain protein 1 (MUPP1) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 9 of MUPP1. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, PDZ9, and PDZ13. This MuPP1-like PDZ9 domain is therefore absent from PATJ. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ9 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467263 [Multi-domain] Cd Length: 79 Bit Score: 55.43 E-value: 1.64e-09
|
||||||||
| PDZ1_MUPP1-like | cd06689 | PDZ domain 1 of multi-PDZ-domain protein 1 (MUPP1) and PATJ (protein-associated tight junction) ... |
420-502 | 1.84e-09 | ||||
PDZ domain 1 of multi-PDZ-domain protein 1 (MUPP1) and PATJ (protein-associated tight junction), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of MUPP1 and PATJ, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467176 [Multi-domain] Cd Length: 102 Bit Score: 56.10 E-value: 1.84e-09
|
||||||||
| PDZ4_Scribble-like | cd06701 | PDZ domain 4 of Drosophila Scribble, human Scribble homolog, and related domains; PDZ (PSD-95 ... |
441-500 | 2.19e-09 | ||||
PDZ domain 4 of Drosophila Scribble, human Scribble homolog, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 4 of Drosophila Scribble (also known as LAP4), human Scribble homolog (also known as hScrib, LAP4, CriB1, ScrB1 and Vartul), and related domains. They belong to the LAP family, which describes proteins that contain either one or four PDZ domains and 16 LRRs (leucine-rich repeats) and function in controlling cell shape, size and subcellular protein localization. In Drosophila, the Scribble complex, comprising Scribble, discs large, and lethal giant larvae, plays a role in apico-basal cell polarity, in other forms of polarity, including regulation of the actin cytoskeleton, cell signaling and vesicular trafficking, and in tumor development. Mammalian Scribble is important in many aspects of cancer development. Scribble and its homologs can be downregulated or overexpressed in cancer; they have a role in cancer beyond their function in loss of cell polarity. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Scribble-like family PDZ4 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467185 [Multi-domain] Cd Length: 98 Bit Score: 55.69 E-value: 2.19e-09
|
||||||||
| PDZ1_Par3-like | cd06691 | PDZ domain 1 of partitioning defective 3 (Par3), and related domains; PDZ (PSD-95 ... |
249-341 | 2.98e-09 | ||||
PDZ domain 1 of partitioning defective 3 (Par3), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of Par3 (or PAR3 or Par-3, also known as Atypical PKC isotype-specific-interacting protein, ASIP) and related domains; Drosophila bazooka PDZ1 belongs to a different PDZ family. Par3 is a scaffold protein involved in organizing cell polarity across animals. Par3 binds numerous molecules both for its recruitment to one pole of the cell and for downstream contributions to polarized cell function. It regulates cell polarity by targeting the Par complex proteins Par6 and atypical protein kinase C (aPKC) to specific cortical sites. Physical interactions between Par3 and the Par complex include: Par-3 PDZ domain 1 binding to the Par6 PDZ domain, Par3 PDZ domain 1 and PDZ domain 3 binding the Par6's PDZ-binding motif, and an interaction with an undefined region of aPKC that requires both Par3 PDZ2 and PDZ3. The PDZ domains of Par3 have also been implicated as potential phosphoinositide signaling integrators, since its second PDZ domain binds to phosphoinositides, and the third PDZ interacts with phosphoinositide phosphatase PTEN. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Par3 family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467178 [Multi-domain] Cd Length: 98 Bit Score: 55.31 E-value: 2.98e-09
|
||||||||
| PDZ2_PDZD7-like | cd10834 | PDZ domain 2 of the canonical isoform 1 of PDZ domain containing 7 (PDZD7), and related ... |
259-332 | 3.09e-09 | ||||
PDZ domain 2 of the canonical isoform 1 of PDZ domain containing 7 (PDZD7), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of the long isoform 1 of PDZD7, and related domains. PDZD7 is critical for the organization of Usher syndrome type 2 (USH2) complex. Usher syndrome is the leading cause of hereditary sensory deaf-blindness in humans; USH2 is the most common sub-type. Formation of the USH2 complex is based upon heterodimerization between PDZD7 and whirlin (another PDZ domain-containing protein) and a subsequent dynamic interplay between USH2 proteins via their multiple PDZ domains. The PDZD7 PDZ2 domain binds GPR98 (also known as VLGR1) and usherin (USH2A). PDZD7 and whirlin form heterodimers through their multiple PDZ domains; whirlin and PDZD7 interact with usherin and GPR98 to form an interdependent ankle link complex. PDZD7 also interacts with myosin VIIa. PDZD7 also forms homodimers through its PDZ2 domain. Various isoforms of PDZD7 produced by alternative splicing have been identified; this subgroup includes the second PDZ domain of the canonical isoform of PDZD7- isoform 1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZD7-like family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467270 [Multi-domain] Cd Length: 85 Bit Score: 54.70 E-value: 3.09e-09
|
||||||||
| zf-C3HC4_2 | pfam13923 | Zinc finger, C3HC4 type (RING finger); |
17-53 | 3.72e-09 | ||||
Zinc finger, C3HC4 type (RING finger); Pssm-ID: 404756 [Multi-domain] Cd Length: 40 Bit Score: 53.21 E-value: 3.72e-09
|
||||||||
| PDZ1_LNX1_2-like | cd06677 | PDZ domain 1 of human Ligand of Numb protein X 1 (LNX1) and LNX2, and related domains; PDZ ... |
242-337 | 3.75e-09 | ||||
PDZ domain 1 of human Ligand of Numb protein X 1 (LNX1) and LNX2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of LNX1 (also known as PDZ domain-containing RING finger protein 2, PDZRN2) and LNX2 (also known as PDZ domain-containing RING finger protein 1, PDZRN1), and related domains. LNX1 and LNX2 are Ring (Really Interesting New Gene) finger and PDZ domain-containing E3 ubiquitin ligases that bind to the cell fate determinant protein NUMB and mediate its ubiquitination. LNX1 can ubiquitinate a number of other ligands including PPFIA1, KLHL11, KIF7 and ERC2. LNX1 and LNX2 each have four PDZ domains. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This LNX family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467165 [Multi-domain] Cd Length: 89 Bit Score: 54.56 E-value: 3.75e-09
|
||||||||
| RING-HC_RNF151 | cd16547 | RING finger, HC subclass, found in RING finger protein 151 (RNF151) and similar proteins; ... |
13-57 | 5.44e-09 | ||||
RING finger, HC subclass, found in RING finger protein 151 (RNF151) and similar proteins; RNF151 is a testis-specific RING finger protein that interacts with dysbindin, a synaptic and microtubular protein that binds brain snapin, a SNARE-binding protein that mediates intracellular membrane fusion in both neuronal and non-neuronal cells. Thus, it may be involved in acrosome formation of spermatids by interacting with multiple proteins participating in membrane biogenesis and microtubule organization. RNF151 contains a C3HC4-type RING finger domain, a putative nuclear localization signal (NLS), and a TNF receptor associated factor (TRAF)-type zinc finger domain. Pssm-ID: 438209 [Multi-domain] Cd Length: 49 Bit Score: 52.85 E-value: 5.44e-09
|
||||||||
| PDZ_SHANK1_3-like | cd06746 | PDZ domain of SH3 and multiple ankyrin repeat domains protein 1 (SHANK1), SHANK2, SHANK3, and ... |
249-337 | 6.41e-09 | ||||
PDZ domain of SH3 and multiple ankyrin repeat domains protein 1 (SHANK1), SHANK2, SHANK3, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of SHANK1, SHANK2, SHANK3, and related domains. SHANK family proteins, SHANK1 (also known as somatostatin receptor-interacting protein, SSTR-interacting protein, SSTRIP), SHANK2 (also known as cortactin-binding protein 1, proline-rich synapse-associated protein 1), and SHANK3 (proline-rich synapse-associated protein 2) are synaptic scaffolding proteins which are highly enriched in the post-synaptic densities of excitatory synapses. They have been implicated in synaptic transmission, synapse formation, synaptic plasticity, and cytoskeletal remodeling, and are regulators of Cav1 calcium current and CREB target expression. Many protein ligands have been identified for the Shank PDZ domain, such as GKAP (also known as SAPAP), betaPIX (a guanine nucleotide exchange factor used by Rho GTPase family members Rac1 and Cdc42), alpha-latrotoxin, neuroligin, group I metabotropic glutamate receptors (mGluRs), and L-type calcium channels. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This SHANK-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged as beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta- strand F. Pssm-ID: 467228 [Multi-domain] Cd Length: 101 Bit Score: 54.52 E-value: 6.41e-09
|
||||||||
| PDZ2_Par3-like | cd23058 | PDZ domain 2 of partitioning defective 3 (Par3), and related domains; PDZ (PSD-95 ... |
246-337 | 6.61e-09 | ||||
PDZ domain 2 of partitioning defective 3 (Par3), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of Par3 (or PAR3 or Par-3, also known as Atypical PKC isotype-specific-interacting protein, ASIP, Drosophila Bazooka) and related domains. Par3 is a scaffold protein involved in organizing cell polarity across animals. Par3 binds numerous molecules both for its recruitment to one pole of the cell and for downstream contributions to polarized cell function. It regulates cell polarity by targeting the Par complex proteins Par6 and atypical protein kinase C (aPKC) to specific cortical sites. Physical interactions between Par3 and the Par complex include Par3 PDZ domain 1 binding to the Par6 PDZ domain, Par3 PDZ domain 1 and PDZ domain 3 binding the Par6's PDZ-binding motif, and an interaction with an undefined region of aPKC that requires both Par3 PDZ2 and PDZ3. The PDZ domains of Par3 have also been implicated as potential phosphoinositide signaling integrators, since its second PDZ domain binds to phosphoinositides, and the third PDZ interacts with phosphoinositide phosphatase PTEN. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Par3 family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467271 [Multi-domain] Cd Length: 93 Bit Score: 54.18 E-value: 6.61e-09
|
||||||||
| PDZ3_DLG5-like | cd06767 | PDZ domain 3 of Discs Large 5 (Dlg5) and related domains; PDZ (PSD-95 (Postsynaptic density ... |
243-335 | 6.95e-09 | ||||
PDZ domain 3 of Discs Large 5 (Dlg5) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 3 of Drosophila and mammalian Dlg5, and related domains. Dlg5 is a scaffold protein with multiple conserved functions that are independent of each other in regulating growth, cell polarity, and cell adhesion. It has a coiled-coil domain, 4 PDZ domains and a MAGUK domain (an SH3 domain next to a non-catalytically active guanylate kinase domain). Deregulation of Dlg5 has been implicated in the malignancy of several cancer types. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Dlg5-like family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467248 [Multi-domain] Cd Length: 82 Bit Score: 53.48 E-value: 6.95e-09
|
||||||||
| PDZ_SYNJ2BP-like | cd06709 | PDZ domain of synaptojanin-2-binding protein (SYNJ2BP), and related domains; PDZ (PSD-95 ... |
443-487 | 7.40e-09 | ||||
PDZ domain of synaptojanin-2-binding protein (SYNJ2BP), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of SYNJ2BP, and related domains. SYNJ2BP (also known as mitochondrial outer membrane protein 25, OMP25) regulates endocytosis of activin type 2 receptor kinases through the Ral/RALBP1-dependent pathway and may be involved in suppression of activin-induced signal transduction. Binding partners of the SYNJ2BP PDZ domain include activin type II receptors (ActR-II), and SYNJ2. SYNJ2BP interacts with the PDZ binding motif of the Notch Delta-like ligand 1 (DLL1) and DLL4, promoting Delta-Notch signaling, and inhibiting sprouting angiogenesis. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This SYNJ2BP-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467193 [Multi-domain] Cd Length: 86 Bit Score: 53.84 E-value: 7.40e-09
|
||||||||
| cpPDZ2_DegP-like | cd23084 | circularly permuted second PDZ domain (PDZ2) of Escherichia coli periplasmic serine ... |
428-502 | 7.49e-09 | ||||
circularly permuted second PDZ domain (PDZ2) of Escherichia coli periplasmic serine endoprotease DegP and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of Escherichia coli DegP (also known as heat shock protein DegP and Protease Do), and related domains. DegP belongs to the HtrA family of housekeeping proteases. It acts as a protease, degrading transiently denatured and unfolded or misfolded proteins which accumulate in the periplasm following heat shock or other stress conditions, and as a molecular chaperone at low temperatures. DegP has two PDZ domains in addition to the protease domain; its PDZ1 domain is responsible for the identifying the distinct substrate sequences that affect degradation (degron) of the substrate sequence, and its PDZ2 domain is responsible for the combining with other DegP monomers to form a stable oligomer structure. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains and as well as those with circular permutations and domain swapping of beta-strands. The canonical PDZ domain contains six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. This DegP family PDZ domain 2 is a circularly permuted PDZ domain which places beta-strand A on the C-terminus. Another permutation exists in the PDZ superfamily which places both beta-strands A and B on the C-terminus. Pssm-ID: 467631 [Multi-domain] Cd Length: 83 Bit Score: 53.40 E-value: 7.49e-09
|
||||||||
| RING-HC_TRIM25_C-IV | cd16597 | RING finger, HC subclass, found in tripartite motif-containing protein TRIM25 and similar ... |
15-66 | 7.60e-09 | ||||
RING finger, HC subclass, found in tripartite motif-containing protein TRIM25 and similar proteins; TRIM25, also known as estrogen-responsive finger protein (EFP), RING finger protein 147 (RNF147), or RING-type E3 ubiquitin transferase, is an E3 ubiquitin/ISG15 ligase that is induced by estrogen and is therefore particularly abundant in placenta and uterus. TRIM25 regulates various cellular processes through E3 ubiquitin ligase activity, transferring ubiquitin and ISG15 to target proteins. It mediates K63-linked polyubiquitination of retinoic acid inducible gene I (RIG-I) that is crucial for downstream antiviral interferon signaling. It is also required for melanoma differentiation-associated gene 5 (MDA5) and mitochondrial antiviral signaling (MAVS, also known as IPS-1, VISA, Cardiff) mediated activation of nuclear factor-kappaB (NF-kappaB) and interferon production. Upon UV irradiation, TRIM25 interacts with mono-ubiquitinated PCNA and promotes its ISG15 modification (ISGylation), suggesting a crucial role in termination of error-prone translesion DNA synthesis. TRIM25 also functions as a novel regulator of p53 and Mdm2. It enhances p53 and Mdm2 abundance by inhibiting their ubiquitination and degradation in 26S proteasomes. Meanwhile, it inhibits p53's transcriptional activity and dampens the response to DNA damage, and is essential for medaka development and this dependence is rescued by silencing of p53. Moreover, TRIM25 is involved in the host cellular innate immune response against retroviral infection. It interferes with the late stage of feline leukemia virus (FeLV) replication. Furthermore, TRIM25 acts as an oncogene in gastric cancer. Its blockade by RNA interference inhibits migration and invasion of gastric cancer cells through transforming growth factor-beta (TGF-beta) signaling, suggesting it presents a novel target for the detection and treatment of gastric cancer. In addition, TRIM25 acts as an RNA-specific activator for Lin28a/TuT4-mediated uridylation. TRIM25 belongs to the C-IV subclass of TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region, as well as a B30.2/SPRY (SplA and ryanodine receptor) domain positioned C-terminal to the RBCC domain. Pssm-ID: 438259 [Multi-domain] Cd Length: 71 Bit Score: 53.08 E-value: 7.60e-09
|
||||||||
| mRING-HC-C3HC3D_LNX1-like | cd16637 | Modified RING finger, HC subclass (C3HC3D-type), found in ligand of Numb protein LNX1, LNX2, ... |
15-53 | 8.09e-09 | ||||
Modified RING finger, HC subclass (C3HC3D-type), found in ligand of Numb protein LNX1, LNX2, and similar proteins; The ligand of Numb protein X (LNX) family, also known as PDZ and RING (PDZRN) family, includes LNX1-5, which can interact with Numb, a key regulator of neurogenesis and neuronal differentiation. LNX5 (also known as PDZK4 or PDZRN4L) shows high sequence homology to LNX3 and LNX4, but it lacks the RING domain. LNX1-4 proteins function as E3 ubiquitin ligases and have a unique domain architecture consisting of an N-terminal RING-HC finger for E3 ubiquitin ligase activity and either two or four PDZ domains necessary for substrate-binding. LNX1/LNX2-like proteins contain a modified C3HC3D-type RING-HC finger and four PDZ domains. This model corresponds to the RING finger. Pssm-ID: 438299 [Multi-domain] Cd Length: 42 Bit Score: 52.01 E-value: 8.09e-09
|
||||||||
| RING-HC_PEX10 | cd16527 | RING finger, HC subclass, found in peroxin-10 (PEX10) and similar proteins; PEX10, also known ... |
17-57 | 8.80e-09 | ||||
RING finger, HC subclass, found in peroxin-10 (PEX10) and similar proteins; PEX10, also known as peroxisome biogenesis factor 10, peroxisomal biogenesis factor 10, peroxisome assembly protein 10, or RING finger protein 69 (RNF69), is an integral peroxisomal membrane protein with two transmembrane regions and a C3HC4-type RING-HC finger within its cytoplasmically exposed C-terminus. It plays an essential role in peroxisome assembly, import of target substrates, and recycling or degradation of protein complexes and amino acids. It is an essential component of the spinal locomotor circuit, and thus its mutations may be involved in peroxisomal biogenesis disorders (PBD). Mutations in human PEX10 also result in autosomal recessive ataxia. Moreover, PEX10 functions as an E3-ubiquitin ligase with an E2, UBCH5C. It mono- or poly-ubiquitinates PEX5, a key player in peroxisomal matrix protein import, in a UBC4-dependent manner, to control PEX5 receptor recycling or degradation. It also links the E2 ubiquitin conjugating enzyme PEX4 to the protein import machinery of the peroxisome. Pssm-ID: 438190 [Multi-domain] Cd Length: 52 Bit Score: 52.23 E-value: 8.80e-09
|
||||||||
| PDZ3_PDZD2-PDZ1_hPro-IL-16-like | cd06759 | PDZ domain 3 of PDZ domain containing 2 (PDZD2), PDZ domain 1 of human pro-interleukin-16 ... |
444-487 | 8.81e-09 | ||||
PDZ domain 3 of PDZ domain containing 2 (PDZD2), PDZ domain 1 of human pro-interleukin-16 (isoform 1, 1332 AA), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 3 of PDZD2, also known as KIAA0300, PIN-1, activated in prostate cancer (AIPC) and PDZ domain-containing protein 3 (PDZK3). PDZD2 has seven PDZ domains. PDZD2 is expressed at exceptionally high levels in the pancreas and certain cancer tissues, such as prostate cancer. It promotes the proliferation of insulinoma cells and is upregulated during prostate tumorigenesis. In osteosarcoma (OS), the microRNA miR-363 acts as a tumor suppressor by inhibiting PDZD2. This family also includes the first PDZ domain (PDZ1) of human pro-interleukin-16 (isoform 1, also known as nPro-Il-16; 1332 amino-acid protein). Precursor IL-16 is cleaved to produce pro-IL-16 and mature IL-16 (derived from the C-terminal 121 AA). Pro-IL-16 functions as a regulator of T cell growth; mature IL-16 is a CD4 ligand that induces chemotaxis and CD25 expression in CD4+ T cells. IL-16 bioactivity has been closely associated with the progression of several different cancers. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZD2-like family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467240 [Multi-domain] Cd Length: 87 Bit Score: 53.43 E-value: 8.81e-09
|
||||||||
| PDZ1_PTPN13_FRMPD2-like | cd06694 | PDZ domain 1 of protein tyrosine phosphatase non-receptor type 13 (PTPN13),FERM and PDZ ... |
444-503 | 8.99e-09 | ||||
PDZ domain 1 of protein tyrosine phosphatase non-receptor type 13 (PTPN13),FERM and PDZ domain-containing protein 2 (FRMPD2), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of PTPN13 [also known as Fas-associated protein-tyrosine phosphatase 1 (FAP-1), protein-tyrosine phosphatase 1E (PTP-E1), and protein-tyrosine phosphatase (PTPL1)], FRMPD2 (also known as PDZ domain-containing protein 4; PDZ domain-containing protein 5C), and related domains. PTPN13 regulates negative apoptotic signaling and mediates phosphoinositide 3-kinase (PI3K) signaling. PTPN13 has five PDZ domains. Proteins known to interact with PTPN13 PDZ domains include: PLEKHA1 and PLEKHA2 via PTPN13-PDZ domain 1, Fas receptor and thyroid receptor-interacting protein 6 via PTPN13-PDZ domain 2, nerve growth factor receptor and protein kinase N2 via PTPN13-PDZ domain 3, PDZ and LIM domain 4 (PDLIM4) via PTPN13-PDZ domains 2 and 4, and brain calpain-2 via PTPN13-PDZ domains 3, 4 and 5. Calpain-2-mediated PTPN13 fragments may be involved in abnormal tau aggregation and increased risk for Alzheimer's disease. FRMPD2 is localized in the basolateral membranes of polarized epithelial cells and is associated with tight junction formation and immune response; it contains 3 PDZ domains. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PTPN13 family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467180 [Multi-domain] Cd Length: 92 Bit Score: 53.55 E-value: 8.99e-09
|
||||||||
| PDZ4_PTPN13-like | cd06696 | PDZ domain 4 of protein tyrosine phosphatase non-receptor type 13 (PTPN13), and related ... |
431-488 | 9.28e-09 | ||||
PDZ domain 4 of protein tyrosine phosphatase non-receptor type 13 (PTPN13), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 4 of PTPN13 [also known as Fas-associated protein-tyrosine phosphatase 1 (FAP-1), protein-tyrosine phosphatase 1E (PTP-E1), and protein-tyrosine phosphatase (PTPL1)] and related domains. PTPN13 regulates negative apoptotic signaling and mediates phosphoinositide 3-kinase (PI3K) signaling. PTPN13 has five PDZ domains. Proteins known to interact with PTPN13 PDZ domains include: PLEKHA1 and PLEKHA2 via PTPN13-PDZ domain 1, Fas receptor and thyroid receptor-interacting protein 6 via PTPN13-PDZ domain 2, nerve growth factor receptor and protein kinase N2 via PTPN13-PDZ domain 3, PDZ and LIM domain 4 (PDLIM4) via PTPN13-PDZ domains 2 and 4, and brain calpain-2 via PTPN13-PDZ domains 3, 4 and 5. Calpain-2-mediated PTPN13 fragments may be involved in abnormal tau aggregation and increased risk for Alzheimer's disease. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PTPN13 family PDZ4 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467182 [Multi-domain] Cd Length: 85 Bit Score: 53.46 E-value: 9.28e-09
|
||||||||
| PDZ3_Scribble-like | cd06702 | PDZ domain 3 of Drosophila Scribble, human Scribble homolog, and related domains; PDZ (PSD-95 ... |
431-490 | 9.41e-09 | ||||
PDZ domain 3 of Drosophila Scribble, human Scribble homolog, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of Drosophila Scribble (also known as LAP4), human Scribble homolog (also known as hScrib, LAP4, CriB1, ScrB1 and Vartul), and related domains. They belong to the LAP family, which describes proteins that contain either one or four PDZ domains and 16 LRRs (leucine-rich repeats) and function in controlling cell shape, size and subcellular protein localization. In Drosophila, the Scribble complex, comprising Scribble, discs large, and lethal giant larvae, plays a role in apico-basal cell polarity, in other forms of polarity, including regulation of the actin cytoskeleton, cell signaling and vesicular trafficking, and in tumor development. Mammalian Scribble is important in many aspects of cancer development. Scribble and its homologs can be downregulated or overexpressed in cancer; they have a role in cancer beyond their function in loss of cell polarity. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Scribble-like family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467186 [Multi-domain] Cd Length: 89 Bit Score: 53.41 E-value: 9.41e-09
|
||||||||
| PDZ3_Par3-like | cd23059 | PDZ domain 3 of partitioning defective 3 (Par3), and related domains; PDZ (PSD-95 ... |
431-502 | 1.16e-08 | ||||
PDZ domain 3 of partitioning defective 3 (Par3), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 3 of Par3 (or PAR3 or Par-3, also known as Atypical PKC isotype-specific-interacting protein, ASIP, Drosophila Bazooka) and related domains. Par3 is a scaffold protein involved in organizing cell polarity across animals. Par3 binds numerous molecules both for its recruitment to one pole of the cell and for downstream contributions to polarized cell function. It regulates cell polarity by targeting the Par complex proteins Par6 and atypical protein kinase C (aPKC) to specific cortical sites. Physical interactions between Par-3 and the Par complex include Par3 PDZ domain 1 binding to the Par6 PDZ domain, Par3 PDZ domain 1 and PDZ domain 3 binding the Par6's PDZ-binding motif, and an interaction with an undefined region of aPKC that requires both Par3 PDZ2 and PDZ3. The PDZ domains of Par3 have also been implicated as potential phosphoinositide signaling integrators, since its second PDZ domain binds to phosphoinositides, and the third PDZ interacts with phosphoinositide phosphatase PTEN. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Par3 family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467272 [Multi-domain] Cd Length: 103 Bit Score: 53.83 E-value: 1.16e-08
|
||||||||
| RING-HC | cd16449 | HC subclass of RING (RING-HC) finger and its variants; The RING finger is a specialized type ... |
16-53 | 1.19e-08 | ||||
HC subclass of RING (RING-HC) finger and its variants; The RING finger is a specialized type of Zn-finger of 40 to 60 residues that binds two atoms of zinc. It is defined by the "cross-brace" motif that chelates zinc atoms by eight amino acid residues, typically Cys or His, arranged in a characteristic spacing. Canonical RING motifs have been categorized into two major subclasses, RING-HC (C3HC4-type) and RING-H2 (C3H2C3-type), according to their Cys/His content. There are also many variants of RING fingers. Some have a different Cys/His pattern. Some lack a single Cys or His residue at typical Zn ligand positions, especially, the fourth or eighth zinc ligand is prevalently exchanged for an Asp, which can chelate Zn in a RING finger as well. This family corresponds to the HC subclass of RING (RING-HC) fingers that are characterized by containing C3HC4-type canonical RING-HC fingers or noncanonical RING-HC finger variants, including C4C4-, C3HC3D-, C2H2C4-, and C3HC5-type modified RING-HC fingers. The canonical RING-HC finger has been defined as C-X2-C-X(9-39)-C-X(1-3)-H-X(2-3)-C-X2-C-X(4-48)-C-X2-C. It binds two Zn ions in a unique "cross-brace" arrangement, which distinguishes it from tandem zinc fingers and other similar motifs. RING-HC fingers can be found in a group of diverse proteins with a variety of cellular functions, including oncogenesis, development, viral replication, signal transduction, the cell cycle, and apoptosis. Many of them are ubiquitin-protein ligases (E3s) that serve as scaffolds for binding to ubiquitin-conjugating enzymes (E2s, also referred to as ubiquitin carrier proteins or UBCs) in close proximity to substrate proteins, which enables efficient transfer of ubiquitin from E2 to the substrates. Pssm-ID: 438113 [Multi-domain] Cd Length: 41 Bit Score: 51.72 E-value: 1.19e-08
|
||||||||
| PDZ_RapGEF2_RapGEF6-like | cd06755 | PDZ domain of Rap guanine nucleotide exchange factor 2 and Rap guanine nucleotide exchange ... |
254-322 | 1.41e-08 | ||||
PDZ domain of Rap guanine nucleotide exchange factor 2 and Rap guanine nucleotide exchange factor 6, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of Rap guanine nucleotide exchange factor 2 (RapGEF2, also named RA-GEF-1, PDZ-GEF1, CNrasGEF and nRapGEP) and Rap guanine nucleotide exchange factor 6 (RapGEF6, also named RA-GEF-2 and PDZ-GEF2). RapGEF2 and RapGEF6 constitute a subfamily of guanine nucleotide exchange factors (GEFs) for RAP small GTPases that is characterized by the possession of the PDZ and Ras/Rap-associating domains. They activate Rap small GTPases, by catalyzing the release of GDP from the inactive GDP-bound forms, thereby accelerating GTP loading to yield the active GTP-bound forms. The PDZ domain of RapGEF6 (also known as PDZ-GEF2) binds junctional adhesion molecule A (JAM-A). PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This RapGEF2 and RapGEF6 family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467237 [Multi-domain] Cd Length: 83 Bit Score: 52.65 E-value: 1.41e-08
|
||||||||
| PDZ2_ZO1-like_ds | cd06728 | PDZ domain 2 of Zonula Occludens-1 (ZO-1), ZO-2 and ZO-3, and related domains; form ... |
420-503 | 1.45e-08 | ||||
PDZ domain 2 of Zonula Occludens-1 (ZO-1), ZO-2 and ZO-3, and related domains; form domain-swapping dimers; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of ZO-1, -2, -3 and related domains. Zonula occludens proteins (ZO-1, ZO-2, ZO-3) are multi-PDZ domain proteins involved in the maintenance and biogenesis of multi-protein networks at the cytoplasmic surface of intercellular contacts in epithelial and endothelial cells. They have three N-terminal PDZ domains, PDZ1-3, followed by a Src homology-3 (SH3) domain and a guanylate kinase (GuK)-like domain. Among protein-protein interactions for all ZO proteins is the binding of the first PDZ domain (PDZ1) to the C-termini of claudins , and the homo- and hetero-dimerization of ZO-proteins via their second PDZ domain (PDZ2), which takes place by symmetrical domain swapping of the first two beta-strands of PDZ2. At the cell level, ZO-1 and ZO-2 are involved in polarity maintenance, gene transcription, cell proliferation, and tumor cell metastasis. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This ZO family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467210 [Multi-domain] Cd Length: 79 Bit Score: 52.61 E-value: 1.45e-08
|
||||||||
| RING-HC_LONFs_rpt2 | cd16514 | second RING finger, HC subclass, found in the LON peptidase N-terminal domain and RING finger ... |
15-60 | 1.56e-08 | ||||
second RING finger, HC subclass, found in the LON peptidase N-terminal domain and RING finger protein family; The LON peptidase N-terminal domain and RING finger protein family includes LONRF1 (also known as RING finger protein 191 or RNF191), LONRF2 (also known as RING finger protein 192, RNF192, or neuroblastoma apoptosis-related protease), LONRF3 (also known as RING finger protein 127 or RNF127), which are characterized by containing two C3HC4-type RING-HC fingers, four tetratricopeptide (TPR) repeats, and an ATP-dependent protease La (LON) substrate-binding domain at the C-terminus. Their biological functions remain unclear. This model corresponds to the second RING-HC finger. Pssm-ID: 438177 [Multi-domain] Cd Length: 45 Bit Score: 51.50 E-value: 1.56e-08
|
||||||||
| PDZ2_DLG5-like | cd06765 | PDZ domain 2 of Discs Large 5 (Dlg5) and related domains; PDZ (PSD-95 (Postsynaptic density ... |
446-490 | 1.56e-08 | ||||
PDZ domain 2 of Discs Large 5 (Dlg5) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of Drosophila and mammalian Dlg5, and related domains. Dlg5 is a scaffold protein with multiple conserved functions that are independent of each other in regulating growth, cell polarity, and cell adhesion. It has a coiled-coil domain, 4 PDZ domains and a MAGUK domain (an SH3 domain next to a non-catalytically active guanylate kinase domain). Deregulation of Dlg5 has been implicated in the malignancy of several cancer types. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Dlg5-like family PSZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467246 [Multi-domain] Cd Length: 77 Bit Score: 52.35 E-value: 1.56e-08
|
||||||||
| PDZ11_MUPP1-PDZ9_PATJ-like | cd06674 | PDZ domain 11 of MUPP1 of multi-PDZ-domain protein 1 (MUPP1), domain 9 of PATJ ... |
248-337 | 1.58e-08 | ||||
PDZ domain 11 of MUPP1 of multi-PDZ-domain protein 1 (MUPP1), domain 9 of PATJ (protein-associated tight junction) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 11 of MUPP1, PDZ domain 9 of PATJ, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ11 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467162 [Multi-domain] Cd Length: 87 Bit Score: 52.67 E-value: 1.58e-08
|
||||||||
| RING-HC_ScPSH1-like | cd16568 | RING finger, HC subclass, found in Saccharomyces cerevisiae POB3/SPT16 histone-associated ... |
16-61 | 1.82e-08 | ||||
RING finger, HC subclass, found in Saccharomyces cerevisiae POB3/SPT16 histone-associated protein 1 (ScPSH1) and similar proteins; ScPSH1 is a Cse4-specific E3 ubiquitin ligase that interacts with the kinetochore protein Pat1 and targets the degradation of budding yeast centromeric histone H3 variant, CENP-ACse4, which is essential for faithful chromosome segregation. ScPSH1 contains a C3HC4-type RING-HC finger and a DNA directed RNA polymerase domain. Pssm-ID: 438230 [Multi-domain] Cd Length: 54 Bit Score: 51.60 E-value: 1.82e-08
|
||||||||
| PDZ1_harmonin | cd06737 | PDZ domain 1 of harmonin isoforms a, b, and c, and related domains; PDZ (PSD-95 (Postsynaptic ... |
254-335 | 1.92e-08 | ||||
PDZ domain 1 of harmonin isoforms a, b, and c, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of harmonin isoforms a, b, and c, and related domains. Harmonin (also known as Usher Type 1C, PDZ-73 and AIE-75) is a key organizer of the Usher (USH) protein interactome. USH syndrome is the leading cause of hereditary sensory deaf-blindness in humans; three clinically distinct types of USH have been identified, type 1 to 3. The gene encoding harmonin (USH1C) is the causative gene for the USH type 1C phenotype. There are at least 10 alternatively spliced isoforms of harmonin, which are divided into three subclasses (a, b, and c). All isoforms contain the first two PDZ domains and the first coiled-coil domain. The a and b isoforms all have a third PDZ domain. The different PDZ domains are responsible for interactions with all known Usher syndrome type 1 proteins, and most Usher syndrome type 2 proteins. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This harmonin family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467219 [Multi-domain] Cd Length: 85 Bit Score: 52.65 E-value: 1.92e-08
|
||||||||
| PDZ4_PTPN13-like | cd06696 | PDZ domain 4 of protein tyrosine phosphatase non-receptor type 13 (PTPN13), and related ... |
255-342 | 1.93e-08 | ||||
PDZ domain 4 of protein tyrosine phosphatase non-receptor type 13 (PTPN13), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 4 of PTPN13 [also known as Fas-associated protein-tyrosine phosphatase 1 (FAP-1), protein-tyrosine phosphatase 1E (PTP-E1), and protein-tyrosine phosphatase (PTPL1)] and related domains. PTPN13 regulates negative apoptotic signaling and mediates phosphoinositide 3-kinase (PI3K) signaling. PTPN13 has five PDZ domains. Proteins known to interact with PTPN13 PDZ domains include: PLEKHA1 and PLEKHA2 via PTPN13-PDZ domain 1, Fas receptor and thyroid receptor-interacting protein 6 via PTPN13-PDZ domain 2, nerve growth factor receptor and protein kinase N2 via PTPN13-PDZ domain 3, PDZ and LIM domain 4 (PDLIM4) via PTPN13-PDZ domains 2 and 4, and brain calpain-2 via PTPN13-PDZ domains 3, 4 and 5. Calpain-2-mediated PTPN13 fragments may be involved in abnormal tau aggregation and increased risk for Alzheimer's disease. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PTPN13 family PDZ4 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467182 [Multi-domain] Cd Length: 85 Bit Score: 52.31 E-value: 1.93e-08
|
||||||||
| PDZ1_Par3-like | cd06691 | PDZ domain 1 of partitioning defective 3 (Par3), and related domains; PDZ (PSD-95 ... |
426-481 | 2.05e-08 | ||||
PDZ domain 1 of partitioning defective 3 (Par3), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of Par3 (or PAR3 or Par-3, also known as Atypical PKC isotype-specific-interacting protein, ASIP) and related domains; Drosophila bazooka PDZ1 belongs to a different PDZ family. Par3 is a scaffold protein involved in organizing cell polarity across animals. Par3 binds numerous molecules both for its recruitment to one pole of the cell and for downstream contributions to polarized cell function. It regulates cell polarity by targeting the Par complex proteins Par6 and atypical protein kinase C (aPKC) to specific cortical sites. Physical interactions between Par3 and the Par complex include: Par-3 PDZ domain 1 binding to the Par6 PDZ domain, Par3 PDZ domain 1 and PDZ domain 3 binding the Par6's PDZ-binding motif, and an interaction with an undefined region of aPKC that requires both Par3 PDZ2 and PDZ3. The PDZ domains of Par3 have also been implicated as potential phosphoinositide signaling integrators, since its second PDZ domain binds to phosphoinositides, and the third PDZ interacts with phosphoinositide phosphatase PTEN. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Par3 family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467178 [Multi-domain] Cd Length: 98 Bit Score: 52.62 E-value: 2.05e-08
|
||||||||
| PDZ3_MUPP1-like | cd06791 | PDZ domain 3 of multi-PDZ-domain protein 1 (MUPP1) and PATJ (protein-associated tight junction) ... |
420-488 | 2.17e-08 | ||||
PDZ domain 3 of multi-PDZ-domain protein 1 (MUPP1) and PATJ (protein-associated tight junction) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 3 of MUPP1 and PATJ, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467253 [Multi-domain] Cd Length: 89 Bit Score: 52.62 E-value: 2.17e-08
|
||||||||
| PDZ_Radil-like | cd06690 | PDZ domain of Ras-associating and dilute domain-containing protein (Radil) and related domains; ... |
248-331 | 2.42e-08 | ||||
PDZ domain of Ras-associating and dilute domain-containing protein (Radil) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of Radil (also known as protein KIAA1849) and related domains. Radil is required for cell adhesion and migration of neural crest precursors during development. Radil is a component of a Rasip1-Radil-ARHGAP29 complex at endothelial cell-cell junctions. Rap1, via its effectors Radil and Rasip1 and their binding partner ArhGAP29, controls the endothelial barrier by decreasing Rho-mediated radial tension on cell-cell junctions. ArhGAP29 binds the Radil PDZ domain. The Radil PDZ domain also binds kinesin family protein 14 (KIF14); KIF14 negatively regulates Rap1-mediated inside-out integrin activation by tethering Radil on microtubules. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Radil-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467177 [Multi-domain] Cd Length: 88 Bit Score: 52.29 E-value: 2.42e-08
|
||||||||
| PDZ1_PDZD7-like | cd10833 | PDZ domain 1 of the canonical isoform 1 of PDZ domain containing 7 (PDZD7), and related ... |
246-321 | 2.89e-08 | ||||
PDZ domain 1 of the canonical isoform 1 of PDZ domain containing 7 (PDZD7), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of the long isoform 1 of PDZD7, and related domains. PDZD7 is critical for the organization of Usher syndrome type 2 (USH2) complex. Usher syndrome is the leading cause of hereditary sensory deaf-blindness in humans; USH2 is the most common sub-type. Formation of the USH2 complex is based upon heterodimerization between PDZD7 and whirlin (another PDZ domain-containing protein) and a subsequent dynamic interplay between USH2 proteins via their multiple PDZ domains. The PDZD7 PDZ2 domain binds GPR98 (also known as VLGR1) and usherin (USH2A). PDZD7 and whirlin form heterodimers through their multiple PDZ domains; whirlin and PDZD7 interact with usherin and GPR98 to form an interdependent ankle link complex. PDZD7 also interacts with myosin VIIa. PDZD7 also forms homodimers through its PDZ2 domain. Various isoforms of PDZD7 produced by alternative splicing have been identified; this subgroup includes the first PDZ domain of the canonical isoform of PDZD7- isoform 1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZD7-like family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467269 [Multi-domain] Cd Length: 84 Bit Score: 52.05 E-value: 2.89e-08
|
||||||||
| PDZ2_DLG5-like | cd06765 | PDZ domain 2 of Discs Large 5 (Dlg5) and related domains; PDZ (PSD-95 (Postsynaptic density ... |
280-340 | 3.07e-08 | ||||
PDZ domain 2 of Discs Large 5 (Dlg5) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of Drosophila and mammalian Dlg5, and related domains. Dlg5 is a scaffold protein with multiple conserved functions that are independent of each other in regulating growth, cell polarity, and cell adhesion. It has a coiled-coil domain, 4 PDZ domains and a MAGUK domain (an SH3 domain next to a non-catalytically active guanylate kinase domain). Deregulation of Dlg5 has been implicated in the malignancy of several cancer types. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Dlg5-like family PSZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467246 [Multi-domain] Cd Length: 77 Bit Score: 51.58 E-value: 3.07e-08
|
||||||||
| PDZ_NHERF-like | cd06768 | PDZ domains of the Na+/H+ exchange regulatory cofactor (NHERF) family (NHERF1-4), and related ... |
251-336 | 3.73e-08 | ||||
PDZ domains of the Na+/H+ exchange regulatory cofactor (NHERF) family (NHERF1-4), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of the Na+/H+ exchange regulatory cofactor (NHERF) family of multi-PDZ-domain-containing scaffolding proteins (NHERF1-4), and related domains. The NHERF family includes NHERF1 (also known as EBP50), NHERF2 (also known as E3KARP; TKA-1; SIP-1), NHERF3 (also known as CAP70; CLAMP; Napi-Cap-1; PDZD1) and NHERF4 (also known as IKEPP; PDZK2; Napi-Cap-2). NHERF1 and NHERF2 have tandem PDZ domains (PDZ1-2); NHERF3 and NHERF4 have four PDZ domains (PDZ1-4). NHERFs are involved in the regulation of multiple receptors or transporters, such as type II sodium-phosphate cotransporter (Npt2a), purinergic P2Y1 receptor P2Y1R, the beta2-adrenergic receptor (beta2-AR), parathyroid hormone receptor type 1 (PTHR), the lysophosphatidic acid receptors (LPARs), sodium-hydrogen exchanger 3 (NHE3), and cystic fibrosis transmembrane conductance regulator (CFTR). NHERF-PDZ1 domain interaction partners include Npt2a, purinergic P2Y1 receptor, beta2-AR, CFTR, PTHR, NH3, G-protein-coupled receptor kinase 6 (GRK6A), platelet-derived growth factor receptor (PDGFR), B1 subunit of the H+ATPase, cholesterol, receptor for activated C-kinase RACK1, aquaporin 9, among others. The NHERF PDZ2 domain interacts with fewer proteins: NHERF1 PDZ2 binds Npt2a, PTHR, beta-catenin, aquaporin 9, and RACK1; NHERF2 PDZ2 binds LPA2, P2Y1R, and NHE3, cGMP-dependent protein kinase type II (cGKII). NHERF4 PDZ1 and PDZ4 bind the epithelial Ca(2+) channels TRPV5 and TRPV6. NHERF2/NHERF3 heterodimerization is mediated by PDZ domains of NHERF2 and the C-terminal PDZ domain recognition motif of NHERF3. NHERF4 regulates several transporters mediating influx of xenobiotics and nutrients in the small intestine. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This NHERF-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467249 [Multi-domain] Cd Length: 80 Bit Score: 51.29 E-value: 3.73e-08
|
||||||||
| PDZ5_PTPN13-like | cd06697 | PDZ domain 5 of protein tyrosine phosphatase non-receptor type 13 (PTPN13), and related ... |
250-337 | 3.93e-08 | ||||
PDZ domain 5 of protein tyrosine phosphatase non-receptor type 13 (PTPN13), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 5 of PTPN13 [also known as Fas-associated protein-tyrosine phosphatase 1 (FAP-1), Protein-tyrosine phosphatase 1E (PTP-E1), and Protein-tyrosine phosphatase (PTPL1)] and related domains. PTPN13 regulates negative apoptotic signaling and mediates phosphoinositide 3-kinase (PI3K) signaling. PTPN13 has five PDZ domains. Proteins known to interact with PTPN13 PDZ domains include: PLEKHA1 and PLEKHA2 via PTPN13-PDZ domain 1, Fas receptor and thyroid receptor-interacting protein 6 via PTPN13-PDZ domain 2, nerve growth factor receptor and protein kinase N2 via PTPN13-PDZ domain 3, PDZ and LIM domain 4 (PDLIM4) via PTPN13-PDZ domains 2 and 4, and brain calpain-2 via PTPN13-PDZ domains 3, 4 and 5. Calpain-2-mediated PTPN13 fragments may be involved in abnormal tau aggregation and increased risk for Alzheimer's disease. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PTPN13 family PDZ5 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467183 [Multi-domain] Cd Length: 87 Bit Score: 51.57 E-value: 3.93e-08
|
||||||||
| PDZ5_MUPP1-like | cd06669 | PDZ domain 5 of multi-PDZ-domain protein 1 (MUPP1), PATJ (protein-associated tight junction) ... |
244-340 | 3.95e-08 | ||||
PDZ domain 5 of multi-PDZ-domain protein 1 (MUPP1), PATJ (protein-associated tight junction) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 5 of MUPP1 and PATJ, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ5 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F Pssm-ID: 467157 [Multi-domain] Cd Length: 98 Bit Score: 51.85 E-value: 3.95e-08
|
||||||||
| PDZ3_MAGI-1_3-like | cd06733 | PDZ domain 3 of membrane-associated guanylate kinase inverted 1 (MAGI-1), MAGI-2, and MAGI-3, ... |
248-322 | 4.82e-08 | ||||
PDZ domain 3 of membrane-associated guanylate kinase inverted 1 (MAGI-1), MAGI-2, and MAGI-3, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 3 of MAGI1, 2, 3 (MAGI is also known as Membrane-associated guanylate kinase, WW and PDZ domain-containing protein) and related domains. MAGI proteins have been implicated in the control of cell migration and invasion through altering the activity of phosphatase and tensin homolog (PTEN) and modulating Akt signaling. Four MAGI proteins have been identified (MAGI1-3 and MAGIX). MAGI1-3 have 6 PDZ domains and bind to the C-terminus of PTEN via their PDZ2 domain. MAGIX has a single PDZ domain that is related to MAGI1-3 PDZ domain 5. Other binding partners for MAGI1 include JAM4, C-terminal tail of high risk HPV-18 E6, megalin, TRAF6, Kir4.1 (basolateral K+ channel subunit), and cadherin 23; for MAGI2, include DASM1, dendrin, axin, beta- and delta-catenin, neuroligin, hyperpolarization-activated cation channels, beta1-adrenergic receptors, NMDA receptor, and TARPs; and for MAGI3 includes LPA2. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MAGI family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as beta-strands A, -B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467215 [Multi-domain] Cd Length: 85 Bit Score: 51.46 E-value: 4.82e-08
|
||||||||
| RING-HC_RNF213 | cd16561 | RING finger, HC subclass, found in RING finger protein 213 (RNF213) and similar proteins; ... |
18-57 | 4.95e-08 | ||||
RING finger, HC subclass, found in RING finger protein 213 (RNF213) and similar proteins; RNF213, also known as ALK lymphoma oligomerization partner on chromosome 17 or Moyamoya steno-occlusive disease-associated AAA+ and RING finger protein (mysterin), is an intracellular soluble protein that functions as an E3 ubiquitin-protein ligase and AAA+ ATPase, which possibly contributes to vascular development through mechanical processes in the cell. It plays a unique role in endothelial cells for proper gene expression in response to inflammatory signals from the environment. Mutations in RNF213 may be associated with Moyamoya disease (MMD), an idiopathic cerebrovascular occlusive disorder prevalent in East Asia. It also acts as a nuclear marker for acanthomorph phylogeny. RNF213 contains two tandem enzymatically active AAA+ ATPase modules and a C3HC4-type RING-HC finger. It can form a huge ring-shaped oligomeric complex. Pssm-ID: 438223 [Multi-domain] Cd Length: 50 Bit Score: 50.35 E-value: 4.95e-08
|
||||||||
| PDZ2_Dlg1-2-4-like | cd06724 | PDZ domain 2 of human discs large homolog 1 (Dlg1), Dlg2, and Dlg4, Drosophila disc large (Dlg) ... |
443-490 | 4.97e-08 | ||||
PDZ domain 2 of human discs large homolog 1 (Dlg1), Dlg2, and Dlg4, Drosophila disc large (Dlg), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of Drosophila Dlg1, human Dlg1,2, and 4 and related domains. Dlg1 (also known as synapse-associated protein Dlg197 or SAP-97), Dlg2 (also known as channel-associated protein of synapse-110, postsynaptic density protein 93, or PSD-93), Dlg4 (also known as postsynaptic density protein 95, PSD-95, synapse-associated protein 90, or SAP-90) each have 3 PDZ domains and belong to the membrane-associated guanylate kinase family. Dlg1 regulates antigen receptor signaling and cell polarity in lymphocytes, B-cell proliferation and antibody production, and TGFalpha bioavailability; its PDZ3 domain binds pro-TGFalpha, and its PDZ2 domain binds the TACE metalloprotease responsible for cleaving pro-TGFalpha to a soluble form. Dlg2 is involved in N-methyl-D-aspartate (NMDA) receptor signaling. It regulates surface expression of NMDA receptors in dorsal horn neurons of the spinal cord, and it also interacts with NMDA receptor subunits and with Shaker-type K+ channel subunits to cluster into a channel complex. Dlg4 PDZ1 domain binds NMDA receptors, and its PDZ2 domain binds neuronal nitric oxide synthase (nNOS), forming a complex in neurons. The Drosophila Scribble complex (Scribble, Dlg, and lethal giant larvae) plays a role in apico-basal cell polarity, and in other forms of polarity, including regulation of the actin cytoskeleton, cell signaling and vesicular trafficking, and in tumor development. Postsynaptic targeting of Drosophila DLG requires interactions mediated by the first two PDZ domains. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Dlg-like family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467207 [Multi-domain] Cd Length: 85 Bit Score: 51.11 E-value: 4.97e-08
|
||||||||
| PDZ1_MUPP1-like | cd06689 | PDZ domain 1 of multi-PDZ-domain protein 1 (MUPP1) and PATJ (protein-associated tight junction) ... |
240-337 | 5.25e-08 | ||||
PDZ domain 1 of multi-PDZ-domain protein 1 (MUPP1) and PATJ (protein-associated tight junction), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of MUPP1 and PATJ, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467176 [Multi-domain] Cd Length: 102 Bit Score: 51.86 E-value: 5.25e-08
|
||||||||
| PDZ1_DLG5-like | cd06764 | PDZ domain 1 of Discs Large 5 (Dlg5) and related domains; PDZ (PSD-95 (Postsynaptic density ... |
244-338 | 5.60e-08 | ||||
PDZ domain 1 of Discs Large 5 (Dlg5) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of Drosophila and mammalian Dlg5, and related domains. Dlg5 is a scaffold protein with multiple conserved functions that are independent of each other in regulating growth, cell polarity, and cell adhesion. It has a coiled-coil domain, 4 PDZ domains and a MAGUK domain (an SH3 domain next to a non-catalytically active guanylate kinase domain). Deregulation of Dlg5 has been implicated in the malignancy of several cancer types. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Dlg5-like family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467245 [Multi-domain] Cd Length: 95 Bit Score: 51.63 E-value: 5.60e-08
|
||||||||
| PDZ2_LNX1_2-like | cd06678 | PDZ domain 2 of human Ligand of Numb protein X 1 (LNX1) and LNX2, and related domains; PDZ ... |
431-502 | 6.50e-08 | ||||
PDZ domain 2 of human Ligand of Numb protein X 1 (LNX1) and LNX2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of LNX1 (also known as PDZ domain-containing RING finger protein 2, PDZRN2) and LNX2 (also known as PDZ domain-containing RING finger protein 1, PDZRN1), and related domains. LNX1 and LNX2 are Ring (Really Interesting New Gene) finger and PDZ domain-containing E3 ubiquitin ligases that bind to the cell fate determinant protein NUMB and mediate its ubiquitination. LNX1 can ubiquitinate a number of other ligands including PPFIA1, KLHL11, KIF7 and ERC2. LNX1 and LNX2 each have four PDZ domains. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This LNX family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467166 [Multi-domain] Cd Length: 82 Bit Score: 50.71 E-value: 6.50e-08
|
||||||||
| PDZ_PDZD11-like | cd06752 | PDZ domain of PDZ domain-containing protein 11, and related domains; PDZ (PSD-95 (Postsynaptic ... |
424-488 | 6.55e-08 | ||||
PDZ domain of PDZ domain-containing protein 11, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of PDZD11, and related domains. PDZD11 (also known as ATPase-interacting PDZ protein, plasma membrane calcium ATPase-interacting single-PDZ protein, PMCA-interacting single-PDZ protein, PISP) is involved in the dynamic assembly of apical junctions (AJs). It is recruited by PLEKHA7 to AJs to promote the efficient junctional recruitment and stabilization of nectins, and the efficient early phases of assembly of AJs in epithelial cells. The PDZD11 PDZ domain binds nectin-1 and nectin-3. PDZD11 also binds to a PDZ binding motif located in the C-terminal tail of the human sodium-dependent multivitamin transporter, to the cytoplasmic tail of the Menkes copper ATPase ATP7A, and to the cytoplasmic tail of all plasma membrane Ca2+-ATPase b-splice variants. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZD11-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467234 [Multi-domain] Cd Length: 83 Bit Score: 50.78 E-value: 6.55e-08
|
||||||||
| PDZ3_harmonin | cd06739 | PDZ domain 3 of harmonin isoforms a and b, and related domains; PDZ (PSD-95 (Postsynaptic ... |
254-327 | 7.21e-08 | ||||
PDZ domain 3 of harmonin isoforms a and b, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 3 of harmonin isoforms a and b, and related domains. Harmonin (also known as Usher Type 1C, PDZ-73 and AIE-75) is a key organizer of the Usher (USH) protein interactome. USH syndrome is the leading cause of hereditary sensory deaf-blindness in humans; three clinically distinct types of USH have been identified, type 1 to 3. The gene encoding harmonin (USH1C) is the causative gene for the USH type 1C phenotype. There are at least 10 alternatively spliced isoforms of harmonin, which are divided into three subclasses (a, b, and c). All isoforms contain the first two PDZ domains and the first coiled-coil domain. The a and b isoforms all have a third PDZ domain. The different PDZ domains are responsible for interactions with all known Usher syndrome type 1 proteins, and most Usher syndrome type 2 proteins. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This harmonin family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467221 [Multi-domain] Cd Length: 94 Bit Score: 51.16 E-value: 7.21e-08
|
||||||||
| PDZ4_INAD-like | cd23065 | PDZ domain 4 of inactivation-no-after-potential D (INAD), and related domains; PDZ (PSD-95 ... |
431-479 | 7.37e-08 | ||||
PDZ domain 4 of inactivation-no-after-potential D (INAD), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 4 of INAD, and related domains. INAD assembles key enzymes of the Drosophila compound eye photo-transduction pathway into a supramolecular complex, supporting efficient and fast light signaling. It contains 5 PDZ domains arranged in tandem (PDZ1-PDZ5) which independently bind various proteins. INAD PDZ2 binds eye-specific protein kinase C, INAD PDZ3 binds transient receptor potential (TRP) channel, and INAD PDZ4,5 tandem binds NORPA (phospholipase Cbeta, PLCbeta). Mutations of the inaD gene that lead to disruption of each of these interactions impair fly photo signal transduction. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This INAD-like family PDZ4 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467278 [Multi-domain] Cd Length: 82 Bit Score: 50.59 E-value: 7.37e-08
|
||||||||
| PDZ_neurabin-like | cd06790 | PDZ domain of neurabin-1 and neurabin-2, and related domains; PDZ (PSD-95 (Postsynaptic ... |
252-328 | 8.39e-08 | ||||
PDZ domain of neurabin-1 and neurabin-2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of neurabin-1 (also known as protein phosphatase 1 regulatory subunit 9A) and neurabin-2 (also known as spinophilin, and protein phosphatase 1 regulatory subunit 9B), and related domains. Neurabin-1 and neurabin-2 are neuronal scaffolding proteins that play important roles in the regulation of synaptic transmission through their ability to interact with and target protein phosphatase 1 (PP1) to dendritic spines where PP1 dephosphorylates and inactivates glutamate receptors. Neurabin-2 interacts with multiple other synaptic proteins, including synaptic signaling and scaffolding proteins (e.g., GluN1 and SAPAP3) and cytoskeletal proteins (e.g., neurofilament medium polypeptide, NF-M). Neurabin-1 and neurabin-2 also binds F-actin. Other binding partners of neurabin-1 include adenosine A1 receptor (A1R), SAD-1 kinase and 70 kDa ribosomal protein S6 kinase (p70-S6K). This PDZ domain is immediately C-terminal to the PP1 binding domain. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This neurabin-like PDZ domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467252 [Multi-domain] Cd Length: 90 Bit Score: 50.88 E-value: 8.39e-08
|
||||||||
| PDZ2_PDZD2-like | cd06758 | PDZ domain 2 of PDZ domain containing 2 (PDZD2), and related domains; PDZ (PSD-95 ... |
444-503 | 8.43e-08 | ||||
PDZ domain 2 of PDZ domain containing 2 (PDZD2), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of PDZD2, also known as KIAA0300, PIN-1, activated in prostate cancer (AIPC) and PDZ domain-containing protein 3 (PDZK3). PDZD2 has seven PDZ domains, and is expressed at exceptionally high levels in the pancreas and certain cancer tissues such as prostate cancer. It promotes the proliferation of insulinoma cells and is upregulated during prostate tumorigenesis. In osteosarcoma (OS), the microRNA miR-363 acts as a tumor suppressor by inhibiting PDZD2. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZD2-like family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467239 [Multi-domain] Cd Length: 88 Bit Score: 50.81 E-value: 8.43e-08
|
||||||||
| PDZ_AFDN-like | cd06789 | PDZ domain of afadin (AFDN), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95) ... |
249-322 | 9.60e-08 | ||||
PDZ domain of afadin (AFDN), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of afadin (AFDN, also known as ALL1-fused gene from chromosome 6 protein (AF6) and MLLT4), and related domains. AFDN belongs to the adhesion system, probably together with the E-cadherin-catenin system, that plays a role in the organization of homotypic, interneuronal, and heterotypic cell-cell adherens junctions. The AFDN PDZ domain interaction partners include poliovirus receptor-related protein PRR2/nectin, the junctional adhesion molecule (JAM), the breakpoint-cluster-region protein (BCR), connexin36 (Cx36), and a subset of Eph-related receptor tyrosine kinases; it can also bind low molecular weight ligands, in competition with a natural peptide ligand. Other AFDN-binding proteins have been identified. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This AFDN family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467251 [Multi-domain] Cd Length: 89 Bit Score: 50.75 E-value: 9.60e-08
|
||||||||
| PDZ_Lin-7-like | cd06796 | PDZ domain of protein Lin-7 and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), ... |
447-490 | 1.03e-07 | ||||
PDZ domain of protein Lin-7 and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of Lin-7 (also known as LIN-7 or LIN7), and related domains. Lin-7 targets and organize protein complexes to epithelial and synaptic plasma membranes. There are three mammalian Lin-7 homologs: Lin-7A (protein lin-7 homolog A, also known as mammalian lin-seven protein 1 (MALS-1), vertebrate lin-7 homolog 1 (Veli-1), tax interaction protein 33); Lin-7B (also known as MALS-2, Veli-2); and Lin-7C (also known as MALS-3, Veli-3). Lin-7 is involved in localization of the Let-23 growth factor receptor to the basolateral membrane of epithelial cells, in tight junction localization of insulin receptor substrate p53 (IRSp53), in retaining gamma-aminobutyric (GABA) transporter (BGT-1) at the basolateral surface of epithelial cells, and in regulating recruitment of neurotransmitter receptors to the postsynaptic density (PSD). The Lin7 PDZ domain binds Let-23, BGT and beta-catenin, and NMDA (N-methyl-D-aspartate) receptor NR2B. Lin-7 also binds to the PDZ binding motif located in the C-terminal tail of Rhotekin, an effector protein for small GTPase Rho. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Lin-7-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467258 [Multi-domain] Cd Length: 86 Bit Score: 50.51 E-value: 1.03e-07
|
||||||||
| PDZ4_DLG5-like | cd06766 | PDZ domain 4 of Discs Large 5 (Dlg5) and related domains; PDZ (PSD-95 (Postsynaptic density ... |
249-335 | 1.19e-07 | ||||
PDZ domain 4 of Discs Large 5 (Dlg5) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 4 of Drosophila and mammalian Dlg5, and related domains. Dlg5 is a scaffold protein with multiple conserved functions that are independent of each other in regulating growth, cell polarity, and cell adhesion. It has a coiled-coil domain, 4 PDZ domains and a MAGUK domain (an SH3 domain next to a non-catalytically active guanylate kinase domain). Deregulation of Dlg5 has been implicated in the malignancy of several cancer types. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Dlg5-like family PDZ4 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467247 [Multi-domain] Cd Length: 81 Bit Score: 50.08 E-value: 1.19e-07
|
||||||||
| PDZ4_INAD-like | cd23065 | PDZ domain 4 of inactivation-no-after-potential D (INAD), and related domains; PDZ (PSD-95 ... |
249-335 | 1.34e-07 | ||||
PDZ domain 4 of inactivation-no-after-potential D (INAD), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 4 of INAD, and related domains. INAD assembles key enzymes of the Drosophila compound eye photo-transduction pathway into a supramolecular complex, supporting efficient and fast light signaling. It contains 5 PDZ domains arranged in tandem (PDZ1-PDZ5) which independently bind various proteins. INAD PDZ2 binds eye-specific protein kinase C, INAD PDZ3 binds transient receptor potential (TRP) channel, and INAD PDZ4,5 tandem binds NORPA (phospholipase Cbeta, PLCbeta). Mutations of the inaD gene that lead to disruption of each of these interactions impair fly photo signal transduction. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This INAD-like family PDZ4 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467278 [Multi-domain] Cd Length: 82 Bit Score: 50.21 E-value: 1.34e-07
|
||||||||
| mRING_PEX12 | cd16451 | Modified RING finger found in peroxin-12 (PEX12) and similar proteins; PEX12, also known as ... |
18-53 | 1.40e-07 | ||||
Modified RING finger found in peroxin-12 (PEX12) and similar proteins; PEX12, also known as peroxisome assembly protein 12 or peroxisome assembly factor 3 (PAF-3), is a RING finger domain-containing integral membrane peroxin required for protein import into peroxisomes. Mutations in human PEX12 result in the peroxisome deficiency Zellweger syndrome of complementation group III (CG-III), a lethal neurological disorder. PEX12 also functions as an E3-ubiquitin ligase that facilitates the PEX4-dependent monoubiquitination of PEX5, a key player in peroxisomal matrix protein import, to control PEX5 receptor recycling or degradation. PEX12 contains a modified RING finger that lacks the third, fourth, and eighth zinc-binding residues of the consensus RING finger motif, suggesting PEX12 may only bind one zinc ion. Pssm-ID: 438115 [Multi-domain] Cd Length: 54 Bit Score: 49.16 E-value: 1.40e-07
|
||||||||
| PDZ7_MUPP1-PD6_PATJ-like | cd06671 | PDZ domain 7 of multi-PDZ-domain protein 1 (MUPP1), PDZ domain 6 of PATJ (protein-associated ... |
420-486 | 1.78e-07 | ||||
PDZ domain 7 of multi-PDZ-domain protein 1 (MUPP1), PDZ domain 6 of PATJ (protein-associated tight junction) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 7 of MUPP1 and PDZ domain 6 of PATJ, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ7 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467159 [Multi-domain] Cd Length: 96 Bit Score: 50.01 E-value: 1.78e-07
|
||||||||
| PDZ3_LNX1_2-like | cd06679 | PDZ domain 3 of human Ligand of Numb protein X 1 (LNX1) and LNX2, and related domains; PDZ ... |
424-488 | 1.79e-07 | ||||
PDZ domain 3 of human Ligand of Numb protein X 1 (LNX1) and LNX2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 3 of LNX1 (also known as PDZ domain-containing RING finger protein 2, PDZRN2) and LNX2 (also known as PDZ domain-containing RING finger protein 1, PDZRN1), and related domains. LNX1 and LNX2 are Ring (Really Interesting New Gene) finger and PDZ domain-containing E3 ubiquitin ligases that bind to the cell fate determinant protein NUMB and mediate its ubiquitination. LNX1 can ubiquitinate a number of other ligands including PPFIA1, KLHL11, KIF7 and ERC2. LNX1 and LNX2 each have four PDZ domains. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This LNX family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467167 [Multi-domain] Cd Length: 88 Bit Score: 49.94 E-value: 1.79e-07
|
||||||||
| RING-HC_TRIM35_C-IV | cd16599 | RING finger, HC subclass, found in tripartite motif-containing protein 35 (TRIM35) and similar ... |
15-69 | 2.05e-07 | ||||
RING finger, HC subclass, found in tripartite motif-containing protein 35 (TRIM35) and similar proteins; TRIM35, also known as hemopoietic lineage switch protein 5 (HLS5), is a putative hepatocellular carcinoma (HCC) suppressor that inhibits phosphorylation of pyruvate kinase isoform M2 (PKM2), which is involved in aerobic glycolysis of cancer cells and further suppresses the Warburg effect and tumorigenicity in HCC. It also negatively regulates Toll-like receptor 7 (TLR7)- and TLR9-mediated type I interferon production by suppressing the stability of interferon regulatory factor 7 (IRF7). Moreover, TRIM35 regulates erythroid differentiation by modulating globin transcription factor 1 (GATA-1) activity. TRIM35 belongs to the C-IV subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region, as well as a B30.2/SPRY (SplA and ryanodine receptor) domain positioned C-terminal to the RBCC domain. Pssm-ID: 438261 [Multi-domain] Cd Length: 66 Bit Score: 49.00 E-value: 2.05e-07
|
||||||||
| PDZ7_PDZD2-PDZ4_hPro-IL-16-like | cd06763 | PDZ domain 7 of PDZ domain containing 2 (PDZD2), PDZ domain 4 of human pro-interleukin-16 ... |
247-320 | 2.14e-07 | ||||
PDZ domain 7 of PDZ domain containing 2 (PDZD2), PDZ domain 4 of human pro-interleukin-16 (isoform 1, 1332 AA), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 7 of PDZD2, also known as KIAA0300, PIN-1, PAPIN, activated in prostate cancer (AIPC) and PDZ domain-containing protein 3 (PDZK3). PDZD2 has seven PDZ domains. PDZD2 is expressed at exceptionally high levels in the pancreas and certain cancer tissues, such as prostate cancer. It promotes the proliferation of insulinoma cells and is upregulated during prostate tumorigenesis. In osteosarcoma (OS), the microRNA miR-363 acts as a tumor suppressor by inhibiting PDZD2. This family include the PDZ domain of the secreted mature form of human interleukin-16 (IL-16); this is the fourth PDZ domain (PDZ4) of human pro-interleukin-16 (isoform 1, also known as nPro-Il-16). Precursor IL-16 is cleaved to produce pro-IL-16 and C-terminal mature IL-16. Pro-IL-16 functions as a regulator of T cell growth; mature IL-16 is a CD4 ligand that induces chemotaxis and CD25 expression in CD4+ T cells. IL-16 bioactivity has been closely associated with the progression of several different cancers PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZD2-like family PDZ7 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467244 [Multi-domain] Cd Length: 86 Bit Score: 49.53 E-value: 2.14e-07
|
||||||||
| PDZ3_ZO1-like_domain | cd06729 | PDZ domain 3 of Zonula Occludens-1 (ZO-1), homologs ZO-2 and ZO-3, and related domains; PDZ ... |
250-321 | 2.37e-07 | ||||
PDZ domain 3 of Zonula Occludens-1 (ZO-1), homologs ZO-2 and ZO-3, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 3 of ZO-1, -2, -3 and related domains. Zonula occludens proteins (ZO-1, ZO-2, ZO-3) are multi-PDZ domain proteins involved in the maintenance and biogenesis of multi-protein networks at the cytoplasmic surface of intercellular contacts in epithelial and endothelial cells. They have three N-terminal PDZ domains, PDZ1-3, followed by a Src homology-3 (SH3) domain and a guanylate kinase (GuK)-like domain. Among protein-protein interactions for all ZO proteins is the binding of the first PDZ domain (PDZ1) to the C-termini of claudins , and the homo- and hetero-dimerization of ZO-proteins via their second PDZ domain (PDZ2), which takes place by symmetrical domain swapping of the first two beta-strands of PDZ2. At the cell level, ZO-1 and ZO-2 are involved in polarity maintenance, gene transcription, cell proliferation, and tumor cell metastasis. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This ZO family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467211 [Multi-domain] Cd Length: 82 Bit Score: 49.10 E-value: 2.37e-07
|
||||||||
| PDZ_AFDN-like | cd06789 | PDZ domain of afadin (AFDN), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95) ... |
431-489 | 2.45e-07 | ||||
PDZ domain of afadin (AFDN), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of afadin (AFDN, also known as ALL1-fused gene from chromosome 6 protein (AF6) and MLLT4), and related domains. AFDN belongs to the adhesion system, probably together with the E-cadherin-catenin system, that plays a role in the organization of homotypic, interneuronal, and heterotypic cell-cell adherens junctions. The AFDN PDZ domain interaction partners include poliovirus receptor-related protein PRR2/nectin, the junctional adhesion molecule (JAM), the breakpoint-cluster-region protein (BCR), connexin36 (Cx36), and a subset of Eph-related receptor tyrosine kinases; it can also bind low molecular weight ligands, in competition with a natural peptide ligand. Other AFDN-binding proteins have been identified. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This AFDN family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467251 [Multi-domain] Cd Length: 89 Bit Score: 49.59 E-value: 2.45e-07
|
||||||||
| PDZ11_MUPP1-PDZ9_PATJ-like | cd06674 | PDZ domain 11 of MUPP1 of multi-PDZ-domain protein 1 (MUPP1), domain 9 of PATJ ... |
431-502 | 2.62e-07 | ||||
PDZ domain 11 of MUPP1 of multi-PDZ-domain protein 1 (MUPP1), domain 9 of PATJ (protein-associated tight junction) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 11 of MUPP1, PDZ domain 9 of PATJ, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ11 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467162 [Multi-domain] Cd Length: 87 Bit Score: 49.20 E-value: 2.62e-07
|
||||||||
| PDZ8_MUPP1-PDZ7_PATJ-PDZ2_INAD-like | cd06672 | PDZ domain 8 of multi-PDZ-domain protein 1 (MUPP1), PDZ domain 7 of protein-associated tight ... |
441-481 | 3.10e-07 | ||||
PDZ domain 8 of multi-PDZ-domain protein 1 (MUPP1), PDZ domain 7 of protein-associated tight junction (PATJ), PDZ domain 2 of Drosophila melanogaster inactivation-no-after-potential D (INAD), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 8 of MUPP1, PDZ domain 7 of PATJ, and PDZ domain 2 of Drosophila melanogaster INAD, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. INAD assembles key enzymes of the Drosophila compound eye photo-transduction pathway into a supramolecular complex, supporting efficient and fast light signaling. It contains 5 PDZ domains arranged in tandem (PDZ1-PDZ5) which independently bind various proteins. INAD PDZ2 binds eye-specific protein kinase C. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ8 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467160 [Multi-domain] Cd Length: 84 Bit Score: 48.83 E-value: 3.10e-07
|
||||||||
| RING-HC_RNF138 | cd16544 | RING finger, HC subclass, found in RING finger protein 138 (RNF138) and similar proteins; ... |
15-64 | 3.27e-07 | ||||
RING finger, HC subclass, found in RING finger protein 138 (RNF138) and similar proteins; RNF138, also known as Nemo-like kinase-associated RING finger protein (NARF) or NLK-associated RING finger protein, is an E3 ubiquitin-protein ligase that plays an important role in glioma cell proliferation, apoptosis, and cell cycle. It specifically cooperates with the E2 conjugating enzyme E2-25K (Hip-2/UbcH1), regulates the ubiquitylation and degradation of T cell factor/lymphoid enhancer factor (TCF/LEF), and further suppresses Wnt-beta-catenin signaling. RNF138, together with three closely related proteins: RNF114, RNF125 and RNF166, forms a novel family of ubiquitin ligases with a C3HC4-type RING-HC finger, a C2HC-, and two C2H2-type zinc fingers, as well as a ubiquitin interacting motif (UIM). Pssm-ID: 438206 [Multi-domain] Cd Length: 53 Bit Score: 47.78 E-value: 3.27e-07
|
||||||||
| mRING-HC-C3HC3D_Nrdp1 | cd16634 | Modified RING finger, HC subclass (C3HC3D-type), found in neuregulin receptor degradation ... |
15-53 | 3.71e-07 | ||||
Modified RING finger, HC subclass (C3HC3D-type), found in neuregulin receptor degradation protein-1 (Nrdp1) and similar proteins; Nrdp1 (referred to as FLRF in mice), also known as RING finger protein 41 (RNF41), is an E3 ubiquitin-protein ligase that plays a critical role in the regulation of cell growth and apoptosis, inflammation and production of reactive oxygen species (ROS), as well as in doxorubicin (DOX)-induced cardiac injury. It promotes the degradation of the epidermal growth factor receptor (EGFR/ErbB) family member, ErbB3, which is independent of growth factor stimulation. It also promotes M2 macrophage polarization by ubiquitinating and activating transcription factor CCAAT/enhancer-binding protein beta (C/EBPbeta) via Lys-63-linked ubiquitination. Moreover, Nrdp1 interacts with and modulates the activity of Parkin, a causative protein for early onset recessive juvenile parkinsonism (AR-JP). It also interacts with ubiquitin-specific protease 8 (USP8), which is involved in trafficking of various transmembrane proteins. Furthermore, Nrdp1 inhibits basal lysosomal degradation and enhances ectodomain shedding of JAK2-associated cytokine receptors. Its phosphorylation by the kinase Par-1b (also known as MARK2) is required for epithelial cell polarity. Nrdp1 contains an N-terminal modified C3HC3D-type RING-HC finger required for enhancing ErbB3 degradation, a B-box, a coiled-coil domain responsible for Nrdp1 oligomerization, and a C-terminal ErbB3-binding domain. Pssm-ID: 438296 [Multi-domain] Cd Length: 43 Bit Score: 47.42 E-value: 3.71e-07
|
||||||||
| PDZ3_Dlg1-2-4-like | cd06795 | PDZ domain 3 of human discs large homolog 1 (Dlg1), Dlg2, and Dlg4, Drosophila disc large (Dlg) ... |
442-504 | 4.21e-07 | ||||
PDZ domain 3 of human discs large homolog 1 (Dlg1), Dlg2, and Dlg4, Drosophila disc large (Dlg), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 3 of Drosophila Dlg1, human Dlg1, 2, and 4 and related domains. Dlg1 (also known as synapse-associated protein Dlg197; SAP-97), Dlg2 (also known as channel-associated protein of synapse-110; postsynaptic density protein 93, PSD-93), Dlg4 (also known as postsynaptic density protein 95, PSD-95; synapse-associated protein 90, SAP-90) each have 3 PDZ domains and belong to the membrane-associated guanylate kinase family. Dlg1 regulates antigen receptor signaling and cell polarity in lymphocytes, B-cell proliferation and antibody production, and TGFalpha bioavailability; its PDZ3 domain binds pro-TGFalpha, and its PDZ2 domain binds the TACE metalloprotease responsible for cleaving pro-TGFalpha to a soluble form. Dlg2 is involved in N-methyl-D-aspartate (NMDA) receptor signaling, regulating surface expression of NMDA receptors in dorsal horn neurons of the spinal cord; it interacts with NMDA receptor subunits and with Shaker-type K+ channel subunits to cluster into a channel complex. The Dlg4 PDZ1 domain binds NMDA receptors, and its PDZ2 domain binds neuronal nitric oxide synthase (nNOS), forming a complex in neurons. The Drosophila Scribble complex (Scribble, Dlg, and lethal giant larvae) plays a role in apico-basal cell polarity, and in other forms of polarity, including regulation of the actin cytoskeleton, cell signaling and vesicular trafficking, and in tumor development; postsynaptic targeting of Drosophila DLG requires interactions mediated by the first two PDZ domains. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Dlg-like family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467257 [Multi-domain] Cd Length: 91 Bit Score: 48.89 E-value: 4.21e-07
|
||||||||
| CtpA | COG0793 | C-terminal processing protease CtpA/Prc, contains a PDZ domain [Posttranslational modification, ... |
431-503 | 4.26e-07 | ||||
C-terminal processing protease CtpA/Prc, contains a PDZ domain [Posttranslational modification, protein turnover, chaperones]; Pssm-ID: 440556 [Multi-domain] Cd Length: 341 Bit Score: 53.34 E-value: 4.26e-07
|
||||||||
| PDZ_GOPC-like | cd06800 | PDZ domain of Golgi-associated PDZ and coiled-coil motif-containing protein (GOPC), and ... |
249-337 | 4.28e-07 | ||||
PDZ domain of Golgi-associated PDZ and coiled-coil motif-containing protein (GOPC), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of GOPC and related domains. GOPC, also known as PIST (PDZ domain protein interacting specifically with TC10), FIG (fused in glioblastoma), and CAL (CFTR-associated ligand), regulates the trafficking of a wide array of proteins, including small GTPases, receptors, and cell surface molecules such as cadherin 23 and CFTR. It may regulate CFTR chloride currents and acid-sensing ASIC3 currents by modulating cell surface expression of both channels, and may play a role in autophagy. Interaction partners of the GOPC PDZ domains include: FZD5, FZD8, ASIC3, CFTR, MUC3, ARFRP1, Ggamma13, neuroligin, and Stargazin. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This GOPC-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467261 [Multi-domain] Cd Length: 83 Bit Score: 48.52 E-value: 4.28e-07
|
||||||||
| RING-HC_RNFT1-like | cd16532 | RING finger, HC subclass, found in RING finger and transmembrane domain-containing protein ... |
18-57 | 4.65e-07 | ||||
RING finger, HC subclass, found in RING finger and transmembrane domain-containing protein RNFT1, RNFT2, and similar proteins; Both RNFT1 and RNFT2 are multi-pass membrane proteins containing a C3HC4-type RING-HC finger. Their biological roles remain unclear. Pssm-ID: 438194 [Multi-domain] Cd Length: 41 Bit Score: 47.30 E-value: 4.65e-07
|
||||||||
| RING-HC_HLTF | cd16509 | RING finger, HC subclass, found in helicase-like transcription factor (HLTF) and similar ... |
15-65 | 4.66e-07 | ||||
RING finger, HC subclass, found in helicase-like transcription factor (HLTF) and similar proteins; HLTF, also known as DNA-binding protein/plasminogen activator inhibitor 1 regulator, HIP116, RING finger protein 80, SWI/SNF-related matrix-associated actin-dependent regulator of chromatin subfamily A member 3, or sucrose nonfermenting protein 2-like 3, is a yeast RAD5 homolog found in mammals. It has both E3 ubiquitin ligase and DNA helicase activities, and plays a pivotal role in the template-switching pathway of DNA damage tolerance. It is involved in Lys-63-linked poly-ubiquitination of proliferating cell nuclear antigen (PCNA) at Lys-164 and in the regulation of DNA damage tolerance. It shows double-stranded DNA translocase activity with 3'-5' polarity, thereby facilitating regression of the replication fork. HLTF contains an N-terminal HIRAN (HIP116 and RAD5 N-terminal) domain, a SWI/SNF helicase domain that is divided into N- and C-terminal parts by an insertion of a C3HC4-type RING-HC finger involved in the poly-ubiquitination of PCNA. Pssm-ID: 438172 [Multi-domain] Cd Length: 53 Bit Score: 47.68 E-value: 4.66e-07
|
||||||||
| PDZ1_MAGI-1_3-like | cd06731 | PDZ domain 1 of membrane-associated guanylate kinase inverted 1 (MAGI-1), MAGI-2, and MAGI-3, ... |
259-326 | 5.29e-07 | ||||
PDZ domain 1 of membrane-associated guanylate kinase inverted 1 (MAGI-1), MAGI-2, and MAGI-3, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of MAGI1, 2, 3 (MAGI is also known as Membrane-associated guanylate kinase, WW and PDZ domain-containing protein) and related domains. MAGI proteins have been implicated in the control of cell migration and invasion through altering the activity of phosphatase and tensin homolog (PTEN) and modulating Akt signaling. Four MAGI proteins have been identified (MAGI1-3 and MAGIX). MAGI1-3 have 6 PDZ domains and bind to the C-terminus of PTEN via their PDZ2 domain. MAGIX has a single PDZ domain that is related to MAGI1-3 PDZ domain 5. Other binding partners for MAGI1 include JAM4, C-terminal tail of high risk HPV-18 E6, megalin, TRAF6, Kir4.1 (basolateral K+ channel subunit), and cadherin 23; for MAGI2, include DASM1, dendrin, axin, beta- and delta-catenin, neuroligin, hyperpolarization-activated cation channels, beta1-adrenergic receptors, NMDA receptor, and TARPs; and for MAGI3 includes LPA2. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MAGI family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as beta-strands A, -B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467213 [Multi-domain] Cd Length: 85 Bit Score: 48.36 E-value: 5.29e-07
|
||||||||
| zf-RING_2 | pfam13639 | Ring finger domain; |
18-57 | 5.69e-07 | ||||
Ring finger domain; Pssm-ID: 433370 [Multi-domain] Cd Length: 44 Bit Score: 47.02 E-value: 5.69e-07
|
||||||||
| PDZ1_FL-whirlin | cd06740 | PDZ domain 1 of the full-length isoform of whirlin and related domains; PDZ (PSD-95 ... |
420-500 | 6.26e-07 | ||||
PDZ domain 1 of the full-length isoform of whirlin and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of the full-length isoform of whirlin and related domains. Whirlin is an essential protein for developmental pathways in photoreceptor cells of the retina and hair cells of the inner ear. The full-length whirlin isoform has two harmonin N-like domains, three PDZ domains, a proline-rich region, and a PDZ-binding motif. Whirlin isoforms may form different complexes at the periciliary membrane complex (PMC) in photoreceptors, and the stereociliary tip and base in inner ear hair cells. It interacts with ADGRV1 and usherin at the PMC; with SANS and RpgrORF15 at the connecting cilium in photoreceptors; with EPS8, MYO15A, p55, and CASK proteins at the stereociliary tip of inner ear hair cells; and with ADGRV1, usherin, and PDZD7 at the stereociliary base in inner ear hair cells. Mutations in the gene encoding whirlin (WHRN; also known as USH2D and DFNB31), have been found to cause either USH2 subtype (USH2D) or autosomal recessive non-syndromic deafness type 31 (DFNB31). Whirlin is the key protein in the USH2 complex (whirlin, usherin and GPR98) which recruits other USH2 causative proteins at the periciliary membrane in photoreceptors and the ankle link of the stereocilia in hair cells. Whirlin's interaction with espin, another stereociliary protein, may be important for the architecture of the USH2 complex. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This whirlin family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467222 [Multi-domain] Cd Length: 82 Bit Score: 48.13 E-value: 6.26e-07
|
||||||||
| PDZ8_MUPP1-PDZ7_PATJ-PDZ2_INAD-like | cd06672 | PDZ domain 8 of multi-PDZ-domain protein 1 (MUPP1), PDZ domain 7 of protein-associated tight ... |
252-337 | 6.33e-07 | ||||
PDZ domain 8 of multi-PDZ-domain protein 1 (MUPP1), PDZ domain 7 of protein-associated tight junction (PATJ), PDZ domain 2 of Drosophila melanogaster inactivation-no-after-potential D (INAD), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 8 of MUPP1, PDZ domain 7 of PATJ, and PDZ domain 2 of Drosophila melanogaster INAD, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. INAD assembles key enzymes of the Drosophila compound eye photo-transduction pathway into a supramolecular complex, supporting efficient and fast light signaling. It contains 5 PDZ domains arranged in tandem (PDZ1-PDZ5) which independently bind various proteins. INAD PDZ2 binds eye-specific protein kinase C. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ8 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467160 [Multi-domain] Cd Length: 84 Bit Score: 48.06 E-value: 6.33e-07
|
||||||||
| PDZ2_FL-whirlin | cd06741 | PDZ domain 2 of the full-length isoform of whirlin and related domains; PDZ (PSD-95 ... |
430-491 | 6.35e-07 | ||||
PDZ domain 2 of the full-length isoform of whirlin and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of the full-length isoform of whirlin and related domains. Whirlin is an essential protein for developmental pathways in photoreceptor cells of the retina and hair cells of the inner ear. The full-length whirlin isoform has two harmonin N-like domains, three PDZ domains, a proline-rich region, and a PDZ-binding motif. Whirlin isoforms may form different complexes at the periciliary membrane complex (PMC) in photoreceptors, and the stereociliary tip and base in inner ear hair cells. It interacts with ADGRV1 and usherin at the PMC; with SANS and RpgrORF15 at the connecting cilium in photoreceptors; with EPS8, MYO15A, p55, and CASK proteins at the stereociliary tip of inner ear hair cells; and with ADGRV1, usherin, and PDZD7 at the stereociliary base in inner ear hair cells. Mutations in the gene encoding whirlin (WHRN; also known as USH2D and DFNB31), have been found to cause either USH2 subtype (USH2D) or autosomal recessive non-syndromic deafness type 31 (DFNB31). Whirlin is the key protein in the USH2 complex (whirlin, usherin and GPR98) which recruits other USH2 causative proteins at the periciliary membrane in photoreceptors and the ankle link of the stereocilia in hair cells. Whirlin's interaction with espin, another stereociliary protein, may be important for the architecture of the USH2 complex. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This whirlin family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467223 [Multi-domain] Cd Length: 84 Bit Score: 48.03 E-value: 6.35e-07
|
||||||||
| RING-H2_DZIP3 | cd16460 | RING finger, H2 subclass, found in DAZ (deleted in azoospermia)-interacting protein 3 (DZIP3) ... |
18-58 | 7.18e-07 | ||||
RING finger, H2 subclass, found in DAZ (deleted in azoospermia)-interacting protein 3 (DZIP3) and similar proteins; DZIP3, also known as RNA-binding ubiquitin ligase of 138 kDa (RUL138) or 2A-HUB protein, is an RNA-binding E3 ubiquitin-protein ligase that interacts with coactivator-associated arginine methyltransferase 1 (CARM1) and acts as a transcriptional coactivator of estrogen receptor (ER) alpha. It is also a histone H2A ubiquitin ligase that catalyzes monoubiquitination of H2A at lysine 119, functioning as a combinatorial component of the repression machinery required for repressing a specific chemokine gene expression program, critically modulating migratory responses to Toll-like receptors (TLR) activation. DZIP3 contains a C3H2C3-type RING-H2 finger at the C-terminus. Pssm-ID: 438123 [Multi-domain] Cd Length: 47 Bit Score: 46.77 E-value: 7.18e-07
|
||||||||
| PDZ_Radil-like | cd06690 | PDZ domain of Ras-associating and dilute domain-containing protein (Radil) and related domains; ... |
446-476 | 7.46e-07 | ||||
PDZ domain of Ras-associating and dilute domain-containing protein (Radil) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of Radil (also known as protein KIAA1849) and related domains. Radil is required for cell adhesion and migration of neural crest precursors during development. Radil is a component of a Rasip1-Radil-ARHGAP29 complex at endothelial cell-cell junctions. Rap1, via its effectors Radil and Rasip1 and their binding partner ArhGAP29, controls the endothelial barrier by decreasing Rho-mediated radial tension on cell-cell junctions. ArhGAP29 binds the Radil PDZ domain. The Radil PDZ domain also binds kinesin family protein 14 (KIF14); KIF14 negatively regulates Rap1-mediated inside-out integrin activation by tethering Radil on microtubules. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Radil-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467177 [Multi-domain] Cd Length: 88 Bit Score: 48.06 E-value: 7.46e-07
|
||||||||
| RING | smart00184 | Ring finger; E3 ubiquitin-protein ligase activity is intrinsic to the RING domain of c-Cbl and ... |
18-53 | 9.24e-07 | ||||
Ring finger; E3 ubiquitin-protein ligase activity is intrinsic to the RING domain of c-Cbl and is likely to be a general function of this domain; Various RING fingers exhibit binding activity towards E2 ubiquitin-conjugating enzymes (Ubc' s) Pssm-ID: 214546 [Multi-domain] Cd Length: 40 Bit Score: 46.35 E-value: 9.24e-07
|
||||||||
| RING-HC_ORTHRUS_rpt2 | cd23139 | second RING finger, HC subclass, found in Arabidopsis thaliana ORTHRUS and similar proteins; ... |
15-42 | 9.95e-07 | ||||
second RING finger, HC subclass, found in Arabidopsis thaliana ORTHRUS and similar proteins; This subfamily includes Arabidopsis thaliana ORTHRUS 1-5. They are E3 ubiquitin-protein ligases that may participate in CpG methylation-dependent transcriptional regulation and/or epigenetic transcriptional silencing. ORTHRUS 1 mediates ubiquitination with the E2 ubiquitin-conjugating enzymes UBC11, UBC8 and UBC8 homologs (e.g. UBC10, UBC11, UBC28 and UBC29) but not with UBC27, UBC30, UBC32, UBC34 and UBC36. ORTHRUS 2 and 5 mediate ubiquitination with the E2 ubiquitin-conjugating enzyme UBC11. ORTHRUS 1 and 2 promote methylation-mediated gene silencing leading, for example, to early flowering. They can bind to CpG, CpNpG, and CpNpN DNA motifs, with a strong preference for methylated forms, and with highest affinity for CpG substrates. Members of this subfamily contain two typical C3HC4-type RING-HC fingers. This model corresponds to the second one. Pssm-ID: 438501 [Multi-domain] Cd Length: 72 Bit Score: 47.07 E-value: 9.95e-07
|
||||||||
| PDZ2_L-delphilin-like | cd06744 | PDZ domain 2 of delphilin (L-delphilin isoform), and related domains; PDZ (PSD-95 ... |
249-322 | 1.03e-06 | ||||
PDZ domain 2 of delphilin (L-delphilin isoform), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of delphilin (also known as glutamate receptor, ionotropic, delta 2-interacting protein 1, L-delphilin). Delphilin, a postsynaptic protein which it is selectively expressed at cerebellar Purkinje cells, links the glutamate receptor delta 2 subunit (GluRdelta2) with the actin cytoskeleton and various signaling molecules. Two alternatively spliced isoforms of delphilin have been characterized: L-delphilin has two PDZ domains, PDZ1 and PDZ2, and S-delphilin has a single PDZ domain (PDZ2). These two isoforms are differently palmitoylated and may be involved in controlling GluRdelta2 signaling in Purkinje cells. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This delphilin-like family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F Pssm-ID: 467226 [Multi-domain] Cd Length: 75 Bit Score: 47.27 E-value: 1.03e-06
|
||||||||
| Sina | pfam03145 | Seven in absentia protein family; The seven in absentia (sina) gene was first identified in ... |
69-132 | 1.13e-06 | ||||
Seven in absentia protein family; The seven in absentia (sina) gene was first identified in Drosophila. The Drosophila Sina protein is essential for the determination of the R7 pathway in photoreceptor cell development: the loss of functional Sina results in the transformation of the R7 precursor cell to a non- neuronal cell type. The Sina protein contains an N-terminal RING finger domain pfam00097. Through this domain, Sina binds E2 ubiquitin-conjugating enzymes (UbcD1) Sina also interacts with Tramtrack (TTK88) via PHYL. Tramtrack is a transcriptional repressor that blocks photoreceptor determination, while PHYL down-regulates the activity of TTK88. In turn, the activity of PHYL requires the activation of the Sevenless receptor tyrosine kinase, a process essential for R7 determination. It is thought that thus Sina targets TTK88 for degradation, therefore promoting the R7 pathway. Murine and human homologs of Sina have also been identified. The human homolog Siah-1 also binds E2 enzymes (UbcH5) and through a series of physical interactions, targets beta-catenin for ubiquitin degradation. Siah-1 expression is enhanced by p53, itself promoted by DNA damage. Thus this pathway links DNA damage to beta-catenin degradation. Sina proteins, therefore, physically interact with a variety of proteins. The N-terminal RING finger domain that binds ubiquitin conjugating enzymes is described in pfam00097, and does not form part of the alignment for this family. The remainder C-terminal part is involved in interactions with other proteins, and is included in this alignment. In addition to the Drosophila protein and mammalian homologs, whose similarity was noted previously, this family also includes putative homologs from Caenorhabditis elegans, Arabidopsis thaliana. Pssm-ID: 460824 [Multi-domain] Cd Length: 198 Bit Score: 50.29 E-value: 1.13e-06
|
||||||||
| PDZ5_DrPTPN13-like | cd23060 | PDZ domain 5 of Danio rerio tyrosine-protein phosphatase non-receptor type 13 (Ptpn13) and ... |
446-488 | 1.30e-06 | ||||
PDZ domain 5 of Danio rerio tyrosine-protein phosphatase non-receptor type 13 (Ptpn13) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 5 of Danio rerio Ptpn13, and related domains. Protein-tyrosine phosphatases (PTPs) dephosphorylate phosphotyrosyl residues in proteins that are phosphorylated by protein tyrosine kinases (PTKs). Danio rerio Ptpn13 is a classical non-receptor-like PTP. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PTPN13-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467273 [Multi-domain] Cd Length: 80 Bit Score: 46.96 E-value: 1.30e-06
|
||||||||
| PDZ1_LNX1_2-like | cd06677 | PDZ domain 1 of human Ligand of Numb protein X 1 (LNX1) and LNX2, and related domains; PDZ ... |
420-490 | 1.46e-06 | ||||
PDZ domain 1 of human Ligand of Numb protein X 1 (LNX1) and LNX2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of LNX1 (also known as PDZ domain-containing RING finger protein 2, PDZRN2) and LNX2 (also known as PDZ domain-containing RING finger protein 1, PDZRN1), and related domains. LNX1 and LNX2 are Ring (Really Interesting New Gene) finger and PDZ domain-containing E3 ubiquitin ligases that bind to the cell fate determinant protein NUMB and mediate its ubiquitination. LNX1 can ubiquitinate a number of other ligands including PPFIA1, KLHL11, KIF7 and ERC2. LNX1 and LNX2 each have four PDZ domains. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This LNX family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467165 [Multi-domain] Cd Length: 89 Bit Score: 47.24 E-value: 1.46e-06
|
||||||||
| PDZ1_Scribble-like | cd06704 | PDZ domain 1 of Drosophila Scribble, human Scribble homolog, and related domains; PDZ (PSD-95 ... |
437-502 | 1.62e-06 | ||||
PDZ domain 1 of Drosophila Scribble, human Scribble homolog, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of Drosophila Scribble (also known as LAP4), human Scribble homolog (also known as hScrib, LAP4, CriB1, ScrB1 and Vartul), and related domains. They belong to the LAP family, which describes proteins that contain either one or four PDZ domains and 16 LRRs (leucine-rich repeats) and function in controlling cell shape, size and subcellular protein localization. In Drosophila, the Scribble complex, comprising Scribble, discs large, and lethal giant larvae, plays a role in apico-basal cell polarity, in other forms of polarity, including regulation of the actin cytoskeleton, cell signaling and vesicular trafficking, and in tumor development. Mammalian Scribble is important in many aspects of cancer development. Scribble and its homologs can be downregulated or overexpressed in cancer; they have a role in cancer beyond their function in loss of cell polarity. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Scribble-like family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467188 [Multi-domain] Cd Length: 87 Bit Score: 46.89 E-value: 1.62e-06
|
||||||||
| RING-HC_RNF114 | cd16540 | RING finger, HC subclass, found in RING finger protein 114 (RNF114) and similar proteins; ... |
15-60 | 1.63e-06 | ||||
RING finger, HC subclass, found in RING finger protein 114 (RNF114) and similar proteins; RNF114, also known as zinc finger protein 228 (ZNF228) or zinc finger protein 313 (ZNF313), is a p21(WAF1)-targeting ubiquitin E3 ligase that interacts with X-linked inhibitor of apoptosis (XIAP)-associated factor 1 (XAF1) and may play a role in p53-mediated cell-fate decisions. It is involved in the immune response to double-stranded RNA in disease pathogenesis. Moreover, RNF114 interacts with A20 and modulates its ubiquitylation. It negatively regulates nuclear factor-kappaB (NF-kappaB)-dependent transcription and positively regulates T-cell activation. RNF114 may play a putative role in the regulation of immune responses, since it corresponds to a novel psoriasis susceptibility gene, ZNF313. RNF114, together with three closely related proteins: RNF125, RNF138 and RNF166, forms a novel family of ubiquitin ligases with a C3HC4-type RING-HC finger, a C2HC-, and two C2H2-type zinc fingers, as well as a ubiquitin interacting motif (UIM). Pssm-ID: 438202 [Multi-domain] Cd Length: 46 Bit Score: 45.91 E-value: 1.63e-06
|
||||||||
| CtpA | COG0793 | C-terminal processing protease CtpA/Prc, contains a PDZ domain [Posttranslational modification, ... |
242-337 | 1.66e-06 | ||||
C-terminal processing protease CtpA/Prc, contains a PDZ domain [Posttranslational modification, protein turnover, chaperones]; Pssm-ID: 440556 [Multi-domain] Cd Length: 341 Bit Score: 51.41 E-value: 1.66e-06
|
||||||||
| PDZ_RIM-like | cd06714 | PDZ domain of Rab3-interacting molecule 1 (RIM), RIM2, piccolo and related domains; PDZ ... |
253-319 | 1.71e-06 | ||||
PDZ domain of Rab3-interacting molecule 1 (RIM), RIM2, piccolo and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of RIM, RIM2, piccolo and related domains. RIM proteins and Gallus gallus protein piccolo (also called aczonin) are involved in neurotransmitter release at presynaptic active zones, the site of vesicle fusion. A protein complex containing RIM proteins positions synaptic vesicles containing synaptotagmin at the active zone. RIM proteins simultaneously activate docking and priming of synaptic vesicles and recruit Ca2+-channels to active zones, thereby connecting primed synaptic vesicles to Ca2+-channels. RIM binding to vesicular Rab proteins (Rab3 and Rab27 isoforms) mediates vesicle docking; RIM binding to Munc13 activates vesicle priming; RIM binding to the Ca2+-channel, both directly and indirectly via RIM-BP, recruits the Ca2+-channels. The RIM PDZ domain interacts with the C-termini of N- and P/Q-type voltage-gated Ca2+-channels. RIM1, RIM2 and piccolo also participate in regulated exocytosis through binding cAMP-GEFII (cAMP-binding protein-guanidine nucleotide exchange factor II). The piccolo PDZ domain binds cAMP-GEFII. RIM2 also plays a role in dendrite formation by melanocytes. Caenorhabditis elegans RIM (also known as unc-10) may be involved in the regulation of defecation and daumone response. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This RIM-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467198 [Multi-domain] Cd Length: 95 Bit Score: 47.16 E-value: 1.71e-06
|
||||||||
| PDZ_ZASP52-like | cd23068 | PDZ domain of Drosophila melanogaster PDZ and LIM domain protein Zasp52 (also known as Zasp), ... |
249-327 | 1.74e-06 | ||||
PDZ domain of Drosophila melanogaster PDZ and LIM domain protein Zasp52 (also known as Zasp), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of Drosophila melanogaster Zasp52 and related domains. Drosophila melanogaster Zasp52 (also known as Z band alternatively spliced PDZ-motif protein or Zasp) colocalizes with integrins at myotendinous junctions and with alpha-actinin at Z-disks and is required for muscle attachment as well as Z-disk assembly and maintenance. The Zasp52 actin-binding site includes the extended PDZ domain and the ZM region. The Zasp52-PDZ domain is required for myofibril assembly. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Zasp52-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467281 [Multi-domain] Cd Length: 82 Bit Score: 46.75 E-value: 1.74e-06
|
||||||||
| PDZ_NHERF-like | cd06768 | PDZ domains of the Na+/H+ exchange regulatory cofactor (NHERF) family (NHERF1-4), and related ... |
441-494 | 1.94e-06 | ||||
PDZ domains of the Na+/H+ exchange regulatory cofactor (NHERF) family (NHERF1-4), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of the Na+/H+ exchange regulatory cofactor (NHERF) family of multi-PDZ-domain-containing scaffolding proteins (NHERF1-4), and related domains. The NHERF family includes NHERF1 (also known as EBP50), NHERF2 (also known as E3KARP; TKA-1; SIP-1), NHERF3 (also known as CAP70; CLAMP; Napi-Cap-1; PDZD1) and NHERF4 (also known as IKEPP; PDZK2; Napi-Cap-2). NHERF1 and NHERF2 have tandem PDZ domains (PDZ1-2); NHERF3 and NHERF4 have four PDZ domains (PDZ1-4). NHERFs are involved in the regulation of multiple receptors or transporters, such as type II sodium-phosphate cotransporter (Npt2a), purinergic P2Y1 receptor P2Y1R, the beta2-adrenergic receptor (beta2-AR), parathyroid hormone receptor type 1 (PTHR), the lysophosphatidic acid receptors (LPARs), sodium-hydrogen exchanger 3 (NHE3), and cystic fibrosis transmembrane conductance regulator (CFTR). NHERF-PDZ1 domain interaction partners include Npt2a, purinergic P2Y1 receptor, beta2-AR, CFTR, PTHR, NH3, G-protein-coupled receptor kinase 6 (GRK6A), platelet-derived growth factor receptor (PDGFR), B1 subunit of the H+ATPase, cholesterol, receptor for activated C-kinase RACK1, aquaporin 9, among others. The NHERF PDZ2 domain interacts with fewer proteins: NHERF1 PDZ2 binds Npt2a, PTHR, beta-catenin, aquaporin 9, and RACK1; NHERF2 PDZ2 binds LPA2, P2Y1R, and NHE3, cGMP-dependent protein kinase type II (cGKII). NHERF4 PDZ1 and PDZ4 bind the epithelial Ca(2+) channels TRPV5 and TRPV6. NHERF2/NHERF3 heterodimerization is mediated by PDZ domains of NHERF2 and the C-terminal PDZ domain recognition motif of NHERF3. NHERF4 regulates several transporters mediating influx of xenobiotics and nutrients in the small intestine. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This NHERF-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467249 [Multi-domain] Cd Length: 80 Bit Score: 46.66 E-value: 1.94e-06
|
||||||||
| PDZ_ARHGEF11-12-like | cd23069 | PDZ domain of ARHGEF11, ARHGEF12, and related domains; PDZ (PSD-95 (Postsynaptic density ... |
248-325 | 2.00e-06 | ||||
PDZ domain of ARHGEF11, ARHGEF12, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of ARHGEF11, ARHGEF12, and related domains. This subfamily includes the GEFs (guanine exchange factors) ARHGEF11 (Rho guanine nucleotide exchange factor 11, known as PDZ-RhoGEF) and ARHGEF12 (Rho guanine nucleotide exchange factor 12, also known as leukemia-associated RhoGEF). GEFs activate Rho GTPases by promoting GTP binding. ARHGEF11/12 are regulators of G protein signaling (RGS) domain-containing GEFs; the RGS domain mediates their binding to and activation of Galpha (and Gq also in the case of ARHGEF12), in response to G-protein coupled receptor activation. ARHGEF11 and 12 are involved in serum-signaling, and regulate Yes-Associated Protein (YAP1)-dependent transcription. The ARHGEF12 PDZ domain binds plexin-B1 and the receptor tyrosine kinase insulin-like growth factor receptor (IGF-R1) beta-subunit. ARHGEF12 also interacts with glutamate receptor delta-1(GluD1), a postsynaptic organizer of inhibitory synapses in cortical pyramidal neurons. The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This ARHGEF11-12-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467282 [Multi-domain] Cd Length: 76 Bit Score: 46.62 E-value: 2.00e-06
|
||||||||
| cpPDZ_CPP-like | cd06782 | circularly permuted PDZ domain of C-terminal processing peptidase (CPP), a serine protease, ... |
431-504 | 2.27e-06 | ||||
circularly permuted PDZ domain of C-terminal processing peptidase (CPP), a serine protease, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of CPP (also known as tail-specific protease, PRC protein, Protease Re, and Photosystem II D1 protein processing peptidase), and related domains. CPP belongs to the peptidase S41A family. It cleaves a C-terminal 11 residue peptide from the precursor form of penicillin-binding protein 3, and may have a role in protecting bacterium from thermal and osmotic stresses. In the plant chloroplast, the enzyme removes the C-terminal extension of the D1 polypeptide of photosystem II. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains and as well as those with circular permutations and domain swapping of beta-strands. The canonical PDZ domain contains six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. This CPP-like PDZ domain is a circularly permuted PDZ domain which places beta-strand A on the C-terminus. Another permutation exists in the PDZ superfamily which places both beta-strands A and B on the C-terminus. Pssm-ID: 467623 [Multi-domain] Cd Length: 88 Bit Score: 46.71 E-value: 2.27e-06
|
||||||||
| PDZ3_LNX1_2-like | cd06679 | PDZ domain 3 of human Ligand of Numb protein X 1 (LNX1) and LNX2, and related domains; PDZ ... |
249-337 | 2.45e-06 | ||||
PDZ domain 3 of human Ligand of Numb protein X 1 (LNX1) and LNX2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 3 of LNX1 (also known as PDZ domain-containing RING finger protein 2, PDZRN2) and LNX2 (also known as PDZ domain-containing RING finger protein 1, PDZRN1), and related domains. LNX1 and LNX2 are Ring (Really Interesting New Gene) finger and PDZ domain-containing E3 ubiquitin ligases that bind to the cell fate determinant protein NUMB and mediate its ubiquitination. LNX1 can ubiquitinate a number of other ligands including PPFIA1, KLHL11, KIF7 and ERC2. LNX1 and LNX2 each have four PDZ domains. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This LNX family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467167 [Multi-domain] Cd Length: 88 Bit Score: 46.48 E-value: 2.45e-06
|
||||||||
| PDZ2_LNX1_2-like | cd06678 | PDZ domain 2 of human Ligand of Numb protein X 1 (LNX1) and LNX2, and related domains; PDZ ... |
280-337 | 2.49e-06 | ||||
PDZ domain 2 of human Ligand of Numb protein X 1 (LNX1) and LNX2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of LNX1 (also known as PDZ domain-containing RING finger protein 2, PDZRN2) and LNX2 (also known as PDZ domain-containing RING finger protein 1, PDZRN1), and related domains. LNX1 and LNX2 are Ring (Really Interesting New Gene) finger and PDZ domain-containing E3 ubiquitin ligases that bind to the cell fate determinant protein NUMB and mediate its ubiquitination. LNX1 can ubiquitinate a number of other ligands including PPFIA1, KLHL11, KIF7 and ERC2. LNX1 and LNX2 each have four PDZ domains. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This LNX family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467166 [Multi-domain] Cd Length: 82 Bit Score: 46.47 E-value: 2.49e-06
|
||||||||
| mRING-HC-C3HC3D_TRAF6 | cd16643 | Modified RING finger, HC subclass (C3HC3D-type), found in tumor necrosis factor (TNF) ... |
18-53 | 2.51e-06 | ||||
Modified RING finger, HC subclass (C3HC3D-type), found in tumor necrosis factor (TNF) receptor-associated factor 6 (TRAF6) and similar proteins; TRAF6, also known as interleukin-1 signal transducer or RING finger protein 85 (RNF85), is a cytoplasmic adapter protein that mediates signals induced by the tumor necrosis factor receptor (TNFR) superfamily and Toll-like receptor (TLR)/interleukin-1 receptor (IL-1R) family. It functions as a mediator involved in the activation of mitogen-activated protein kinase (MAPK), phosphoinositide 3-kinase (PI3K), and interferon regulatory factor pathways, as well as in IL-1R-mediated activation of NF-kappaB. TRAF6 is also an oncogene that plays a vital role in K-RAS-mediated oncogenesis. TRAF6 contains an N-terminal domain with a modified C3HC3D-type RING-HC finger and several zinc fingers, and a C-terminal TRAF domain that comprises a coiled coil domain and a conserved TRAF-C domain. Pssm-ID: 438305 [Multi-domain] Cd Length: 58 Bit Score: 45.45 E-value: 2.51e-06
|
||||||||
| PEX10 | COG5574 | RING-finger-containing E3 ubiquitin ligase [Posttranslational modification, protein turnover, ... |
15-66 | 2.81e-06 | ||||
RING-finger-containing E3 ubiquitin ligase [Posttranslational modification, protein turnover, chaperones]; Pssm-ID: 227861 [Multi-domain] Cd Length: 271 Bit Score: 50.28 E-value: 2.81e-06
|
||||||||
| PDZ_PTPN3-4-like | cd06706 | PDZ domain of tyrosine-protein phosphatase non-receptor type 3 (PTPN3), tyrosine-protein ... |
250-329 | 2.81e-06 | ||||
PDZ domain of tyrosine-protein phosphatase non-receptor type 3 (PTPN3), tyrosine-protein phosphatase non-receptor type 4 (PTNP4), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of PTPN3, PTPN4 and related domains. PTPN3 (also known as protein-tyrosine phosphatase H1, PTP-H1) has a tumor-suppressive or a tumor-promoting role in many cancers. It serves as a specific phosphatase for the MAP kinase p38gamma; the two interact via their PDZ domains and cooperate to promote Ras-induced oncogenesis. Interaction partners of the PTPN3 PDZ domain include p38gamma and human papillomavirus E6 oncoprotein. PTPN4 (also known as protein-tyrosine phosphatase MEG1) plays a role in immunity, learning, synaptic plasticity or cell homeostasis. p38gamma is also an interaction partner of the PTPN4 PDZ domain: PTPN4 regulates neuronal cell homeostasis by protecting neurons against apoptosis; binding of the C terminus of p38gamma to the PDZ domain of PTPN4, antagonizes the catalytic autoinhibition of PTPN4, leading to cell apoptosis. Other interaction partners of the PTPN4 PDZ domain include glutamate receptor subunit GluN2A, and RABV strain G protein, among others. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PTPN3-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467190 [Multi-domain] Cd Length: 90 Bit Score: 46.54 E-value: 2.81e-06
|
||||||||
| PDZ2_MUPP1-like | cd06667 | PDZ domain 2 of multi-PDZ-domain protein 1 (MUPP1) and PATJ (protein-associated tight junction) ... |
249-339 | 2.92e-06 | ||||
PDZ domain 2 of multi-PDZ-domain protein 1 (MUPP1) and PATJ (protein-associated tight junction) and similar domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of MUPP1 and PATJ, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F Pssm-ID: 467155 [Multi-domain] Cd Length: 80 Bit Score: 46.12 E-value: 2.92e-06
|
||||||||
| RING-HC_TRIM77_C-IV | cd16543 | RING finger, HC subclass, found in tripartite motif-containing protein 77 (TRIM77) and similar ... |
13-57 | 3.14e-06 | ||||
RING finger, HC subclass, found in tripartite motif-containing protein 77 (TRIM77) and similar proteins; TRIM77 belongs to the C-IV subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including two consecutive zinc-binding domains, a C3HC4-type RING-HC finger and Bbox2, as well as a SPRY/B30.2 domain positioned C-terminal to the RBCC domain. Pssm-ID: 438205 [Multi-domain] Cd Length: 54 Bit Score: 45.08 E-value: 3.14e-06
|
||||||||
| RING-HC_CHR27-like | cd23142 | RING finger, HC subclass, found in Arabidopsis thaliana protein CHROMATIN REMODELING 27 (CHR27) ... |
18-65 | 3.32e-06 | ||||
RING finger, HC subclass, found in Arabidopsis thaliana protein CHROMATIN REMODELING 27 (CHR27) and similar proteins; CHR27, also called protein SNF2-RING-HELICASE-LIKE 1, is a probable helicase-like transcription factor involved in transcriptional gene silencing. It associates with SUVR2 and contributes to transcriptional gene silencing at RNA-directed DNA methylation (RdDM) target loci but also at RdDM-independent target loci. It may be involved in nucleosome positioning to form ordered nucleosome arrays on chromatin. It associates with SUVR2 and functions redundantly with FRG2. It is required for the efficient methylation of a broad range of RdDM target loci. CHR27 contains a typical C3HC4-type RING-HC finger. Pssm-ID: 438504 [Multi-domain] Cd Length: 55 Bit Score: 45.25 E-value: 3.32e-06
|
||||||||
| PDZ12_MUPP1-like | cd06675 | PDZ domain 12 of multi-PDZ-domain protein 1 (MUPP1), PDZ domain 10 of protein-associated tight ... |
426-497 | 3.38e-06 | ||||
PDZ domain 12 of multi-PDZ-domain protein 1 (MUPP1), PDZ domain 10 of protein-associated tight junction (PATJ, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 12 of MUPP1, PDZ domain 10 of PATJ, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like PDZ12 family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F Pssm-ID: 467163 [Multi-domain] Cd Length: 86 Bit Score: 46.20 E-value: 3.38e-06
|
||||||||
| PDZ1_PDZD7-like | cd10833 | PDZ domain 1 of the canonical isoform 1 of PDZ domain containing 7 (PDZD7), and related ... |
426-502 | 3.51e-06 | ||||
PDZ domain 1 of the canonical isoform 1 of PDZ domain containing 7 (PDZD7), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of the long isoform 1 of PDZD7, and related domains. PDZD7 is critical for the organization of Usher syndrome type 2 (USH2) complex. Usher syndrome is the leading cause of hereditary sensory deaf-blindness in humans; USH2 is the most common sub-type. Formation of the USH2 complex is based upon heterodimerization between PDZD7 and whirlin (another PDZ domain-containing protein) and a subsequent dynamic interplay between USH2 proteins via their multiple PDZ domains. The PDZD7 PDZ2 domain binds GPR98 (also known as VLGR1) and usherin (USH2A). PDZD7 and whirlin form heterodimers through their multiple PDZ domains; whirlin and PDZD7 interact with usherin and GPR98 to form an interdependent ankle link complex. PDZD7 also interacts with myosin VIIa. PDZD7 also forms homodimers through its PDZ2 domain. Various isoforms of PDZD7 produced by alternative splicing have been identified; this subgroup includes the first PDZ domain of the canonical isoform of PDZD7- isoform 1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZD7-like family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467269 [Multi-domain] Cd Length: 84 Bit Score: 45.89 E-value: 3.51e-06
|
||||||||
| PDZ13_MUPP1-like | cd06676 | PDZ domain 13 of multi-PDZ-domain protein 1 (MUPP1) and related domains; PDZ (PSD-95 ... |
444-488 | 3.98e-06 | ||||
PDZ domain 13 of multi-PDZ-domain protein 1 (MUPP1) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 13 of MUPP1. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, PDZ9, and PDZ13. This MuPP1-like PDZ13 domain is therefore absent from PATJ. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ13 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467164 [Multi-domain] Cd Length: 83 Bit Score: 45.79 E-value: 3.98e-06
|
||||||||
| RING-HC_RNF125 | cd16542 | RING finger, HC subclass, found in RING finger protein 125 (RNF125); RNF125, also known as ... |
15-63 | 4.01e-06 | ||||
RING finger, HC subclass, found in RING finger protein 125 (RNF125); RNF125, also known as T-cell RING activation protein 1 (TRAC-1), is an E3 ubiquitin-protein ligase that is predominantly expressed in lymphoid cells, and functions as a positive regulator of T cell activation. It also down-modulates HIV replication and inhibits pathogen-induced cytokine production. It negatively regulates type I interferon signaling, which conjugates Lys(48)-linked ubiquitination to retinoic acid-inducible gene-I (RIG-I) and subsequently leads to the proteasome-dependent degradation of RIG-I. Further, RNF125 conjugates ubiquitin to melanoma differentiation-associated gene 5 (MDA5), a family protein of RIG-I. It thus acts as a negative regulator of RIG-I signaling, and is a direct target of miR-15b in the context of Japanese encephalitis virus (JEV) infection. Moreover, RNF125 binds to and ubiquitinates JAK1, prompting its degradation and inhibition of receptor tyrosine kinase (RTK) expression. It also negatively regulates p53 function through physical interaction and ubiquitin-mediated proteasome degradation. Mutations in RNF125 may lead to overgrowth syndromes (OGS). RNF125, together with three closely related proteins: RNF114, RNF138 and RNF166, forms a novel family of ubiquitin ligases with a C3HC4-type RING-HC finger, a C2HC-, and two C2H2-type zinc fingers, as well as a ubiquitin interacting motif (UIM). The UIM of RNF125 binds K48-linked poly-ubiquitin chains and is, together with the RING domain, required for auto-ubiquitination. Pssm-ID: 438204 [Multi-domain] Cd Length: 50 Bit Score: 44.87 E-value: 4.01e-06
|
||||||||
| mRING-HC-C3HC3D_LNX2 | cd16780 | Modified RING finger, HC subclass (C3HC3D-type), found in ligand of numb protein X 2 (LNX2); ... |
13-53 | 4.25e-06 | ||||
Modified RING finger, HC subclass (C3HC3D-type), found in ligand of numb protein X 2 (LNX2); LNX2, also known as numb-binding protein 2, or PDZ domain-containing RING finger protein 1 (PDZRN1), is a PDZ domain-containing RING-type E3 ubiquitin ligase responsible for the ubiquitination and degradation of Numb, a component of the Notch signaling pathway that functions in the specification of cell fates during development and is known to control cell numbers during neurogenesis in vertebrates. It interacts with contactin-associated protein 4 (Caspr4, also known as CNTNAP4) in a PDZ domain-dependent manner, which modulates the proliferation and neuronal differentiation of neural progenitor cells (NPCs). LNX2 contains an N-terminal modified C3HC3D-type RING-HC finger, a NPAF motif for Numb/ Numblike-LNX interaction, and four PDZ domains necessary for the binding of substrates, including ErbB2, RhoC, the presynaptic protein CAST, the melanoma/cancer-testis antigen MAGEB18 and several proteins associated with cell junctions, such as JAM4 and the Coxsackievirus and adenovirus receptor (CAR). Pssm-ID: 319694 [Multi-domain] Cd Length: 45 Bit Score: 44.48 E-value: 4.25e-06
|
||||||||
| RING-HC_AtBARD1-like | cd23146 | RING finger, HC subclass, found in Arabidopsis thaliana BRCA1-associated RING domain protein 1 ... |
15-65 | 4.40e-06 | ||||
RING finger, HC subclass, found in Arabidopsis thaliana BRCA1-associated RING domain protein 1 (AtBARD1) and similar proteins; AtBARD1, also called protein REPRESSOR OF WUSCHEL 1, binds specifically to H3K4me3 regions of target gene (e.g. WUS and WOX5) promoters to repress their transcription via chromatin remodeling. It is required for the shoot apical meristem (SAM) organization and maintenance, by confining WUS expression to the organizing center, and for the quiescent center (QC) development in the root apical meristem (RAM), by repressing WOX5 expression in the root proximal meristem. AtBARD1 plays a role in DNA repair and in cell-cycle control. It is required for the repair of DNA double-strand breaks (DSBs), both natural and induced by genotoxic stress, by homologous recombination (HR). AtBARD1 contains a typical C3HC4-type RING-HC finger. Pssm-ID: 438508 [Multi-domain] Cd Length: 54 Bit Score: 44.77 E-value: 4.40e-06
|
||||||||
| RING-HC_TRIM39_C-IV | cd16601 | RING finger, HC subclass, found in tripartite motif-containing protein 39 (TRIM39) and similar ... |
16-45 | 4.40e-06 | ||||
RING finger, HC subclass, found in tripartite motif-containing protein 39 (TRIM39) and similar proteins; TRIM39, also known as RING finger protein 23 (RNF23) or testis-abundant finger protein, is an E3 ubiquitin-protein ligase that plays a role in controlling DNA damage-induced apoptosis through inhibition of the anaphase promoting complex (APC/C), a multiprotein ubiquitin ligase that controls multiple cell cycle regulators, including cyclins, geminin, and others. TRIM39 also functions as a regulator of several key processes in the proliferative cycle. It directly regulates p53 stability. It modulates cell cycle progression and DNA damage responses via stabilizing p21. Moreover, TRIM39 negatively regulates the nuclear factor kappaB (NFkappaB)-mediated signaling pathway through stabilization of Cactin, an inhibitor of NFkappaB- and Toll-like receptor (TLR)-mediated transcription, which is induced by inflammatory stimulants such as tumor necrosis factor alpha. Furthermore, TRIM39 is a MOAP-1-binding protein that can promote apoptosis signaling through stabilization of MOAP-1 via the inhibition of its poly-ubiquitination process. TRIM39 belongs to the C-IV subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region, as well as a B30.2/SPRY (SplA and ryanodine receptor) domain positioned C-terminal to the RBCC domain. Pssm-ID: 438263 [Multi-domain] Cd Length: 44 Bit Score: 44.40 E-value: 4.40e-06
|
||||||||
| cpPDZ_HtrA-like | cd06785 | circularly permuted PDZ domain of high-temperature requirement factor A (HtrA) family serine ... |
278-346 | 4.51e-06 | ||||
circularly permuted PDZ domain of high-temperature requirement factor A (HtrA) family serine proteases and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of HtrA family serine proteases including human HtrA1, HtrA2 (mitochondrial), HtrA3, and HtrA4, and related domains. These proteases are key enzymes associated with pregnancy. Their diverse biological functions include cell growth proliferation, migration and apoptosis. They are also implicated in disorders including Alzheimer's, Parkinson's, arthritis and cancer. HtrA1 (also known as high-temperature requirement A serine peptidase 1, L56, and serine protease 11) substrates include extracellular matrix proteins, proteoglycans, and insulin-like growth factor (IGF)-binding proteins. HtrA1 also inhibits signaling by members of the transforming growth factor beta (TGF-beta) family. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping of beta-strands. The canonical PDZ domain contains six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. This HtrA-like PDZ domain is a circularly permuted PDZ domain which places beta-strand A on the C-terminus. Another permutation exists in the PDZ superfamily which places both beta-strands A and B on the C-terminus. Pssm-ID: 467624 [Multi-domain] Cd Length: 98 Bit Score: 45.95 E-value: 4.51e-06
|
||||||||
| mRING-HC-C3HC3D_LNX1 | cd16779 | Modified RING finger, HC subclass (C3HC3D-type), found in ligand of numb protein X 1 (LNX1); ... |
15-53 | 4.91e-06 | ||||
Modified RING finger, HC subclass (C3HC3D-type), found in ligand of numb protein X 1 (LNX1); LNX1, also known as numb-binding protein 1 or PDZ domain-containing RING finger protein 2, is a PDZ domain-containing RING-type E3 ubiquitin ligase responsible for the ubiquitination and degradation of Numb, a component of the Notch signaling pathway that functions in the specification of cell fates during development and is known to control cell numbers during neurogenesis in vertebrates. LNX1 contains an N-terminal modified C3HC3D-type RING-HC finger, a NPAY motif for Numb-LNX interaction, and four PDZ domains necessary for the binding of substrates, including CAR, ErbB2, SKIP, JAM4, CAST, c-Src, Claudins, RhoC, KCNA4, PAK6, PLEKHG5, PKC-alpha1, TYK2, PDZ-binding kinase (PBK), LNX2, and itself. Pssm-ID: 438435 [Multi-domain] Cd Length: 42 Bit Score: 44.41 E-value: 4.91e-06
|
||||||||
| RING-H2_PA-TM-RING | cd16454 | RING finger, H2 subclass, found in the PA-TM-RING ubiquitin ligase family; The PA-TM-RING ... |
18-57 | 5.76e-06 | ||||
RING finger, H2 subclass, found in the PA-TM-RING ubiquitin ligase family; The PA-TM-RING family represents a group of transmembrane-type E3 ubiquitin ligases, which has been characterized by an N-terminal transient signal peptide, a PA (protease-associated) domain, a TM (transmembrane) domain, as well as a C-terminal C3H2C3-type RING-H2 finger domain. It includes RNF13, RNF167, ZNRF4 (zinc and RING finger 4), GRAIL (gene related to anergy in lymphocytes)/RNF128, RNF130, RNF133, RNF148, RNF149 and RNF150 (which are more closely related), as well as RNF43 and ZNRF3, which have substantially longer C-terminal tail extensions compared with the others. PA-TM-RING proteins are expressed at low levels in all mammalian tissues and species, but they are not present in yeast. They play a common regulatory role in intracellular trafficking/sorting, suggesting that abrogation of their function may result in dysregulation of cellular signaling events in cancer. Pssm-ID: 438118 [Multi-domain] Cd Length: 43 Bit Score: 44.19 E-value: 5.76e-06
|
||||||||
| rad18 | TIGR00599 | DNA repair protein rad18; All proteins in this family for which functions are known are ... |
12-57 | 5.83e-06 | ||||
DNA repair protein rad18; All proteins in this family for which functions are known are involved in nucleotide excision repair.This family is based on the phylogenomic analysis of JA Eisen (1999, Ph.D. Thesis, Stanford University). [DNA metabolism, DNA replication, recombination, and repair] Pssm-ID: 273165 [Multi-domain] Cd Length: 397 Bit Score: 50.00 E-value: 5.83e-06
|
||||||||
| RING-HC_RNF10 | cd16536 | RING finger, HC subclass, found in RING finger protein 10 (RNF10) and similar proteins; RNF10 ... |
17-56 | 6.23e-06 | ||||
RING finger, HC subclass, found in RING finger protein 10 (RNF10) and similar proteins; RNF10 is an E3 ubiquitin-protein ligase that interacts with mesenchyme Homeobox 2 (MEOX2) transcription factor, a regulator of the proliferation, differentiation and migration of vascular smooth muscle cells and cardiomyocytes; it enhances Meox2 activation of the p21 promoter. It also regulates the expression of myelin-associated glycoprotein (MAG) genes and is required for myelin production in Schwann cells of the peripheral nervous system. Moreover, RNF10 regulates retinoic acid-induced neuronal differentiation and the cell cycle exit of P19 embryonic carcinoma cells. RNF10 contains a C3HC4-type RING-HC finger and three putative nuclear localization signals. Pssm-ID: 438198 [Multi-domain] Cd Length: 54 Bit Score: 44.54 E-value: 6.23e-06
|
||||||||
| RING-HC_IRC20-like | cd23135 | RING finger, HC subclass, found in Saccharomyces cerevisiae increased recombination centers ... |
13-53 | 6.31e-06 | ||||
RING finger, HC subclass, found in Saccharomyces cerevisiae increased recombination centers protein 20 (IRC20) and similar proteins; IRC20 is an uncharacterized ATP-dependent helicase that is probably involved in a pathway contributing to genomic integrity. IRC20 contains a typical C3HC4-type RING-HC finger. Pssm-ID: 438497 [Multi-domain] Cd Length: 44 Bit Score: 44.04 E-value: 6.31e-06
|
||||||||
| RING-HC_LONFs_rpt1 | cd16513 | first RING finger, HC subclass, found in the LON peptidase N-terminal domain and RING finger ... |
16-57 | 6.44e-06 | ||||
first RING finger, HC subclass, found in the LON peptidase N-terminal domain and RING finger protein family; The LON peptidase N-terminal domain and RING finger protein family includes LONRF1 (also known as RING finger protein 191 or RNF191), LONRF2 (also known as RING finger protein 192, RNF192, or neuroblastoma apoptosis-related protease), LONRF3 (also known as RING finger protein 127 or RNF127), which are characterized by containing two C3HC4-type RING-HC fingers, four tetratricopeptide (TPR) repeats, and an ATP-dependent protease La (LON) substrate-binding domain at the C-terminus. Their biological functions remain unclear. This model corresponds to the first RING-HC finger. Pssm-ID: 438176 [Multi-domain] Cd Length: 47 Bit Score: 44.22 E-value: 6.44e-06
|
||||||||
| RING-HC_TRIM47-like_C-IV | cd16604 | RING finger, HC subclass, found in tripartite motif-containing protein 47 (TRIM47) and similar ... |
16-57 | 6.56e-06 | ||||
RING finger, HC subclass, found in tripartite motif-containing protein 47 (TRIM47) and similar proteins; TRIM47, also known as gene overexpressed in astrocytoma protein (GOA) or RING finger protein 100 (RNF100), belongs to the C-IV subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, a B-box, and two coiled coil domains, as well as a B30.2/SPRY (SplA and ryanodine receptor) domain positioned C-terminal to the RBCC domain. It plays an important role in the process of dedifferentiation that is associated with astrocytoma tumorigenesis. This subfamily also includes RING finger protein 135 (RNF135). RNF135, also known as RIG-I E3 ubiquitin ligase (REUL) or Riplet, is a widely expressed E3 ubiquitin-protein ligase that consists of an N-terminal C3HC4-type RING-HC finger and C-terminal B30.2/SPRY and PRY motifs, but lacks the B-box and coiled-coil domains that are also typically present in TRIM proteins. RNF135 serves as a specific retinoic acid-inducible gene-I (RIG-I)-interacting protein that ubiquitinates RIG-I and specifically stimulates RIG-I-mediated innate antiviral activity to produce antiviral type-I interferon (IFN) during the early phase of viral infection. It also has been identified as a bio-marker and therapy target of glioblastoma. It associates with the ERK signal transduction pathway and plays a role in glioblastoma cell proliferation, migration and cell cycle. Pssm-ID: 438266 [Multi-domain] Cd Length: 49 Bit Score: 44.33 E-value: 6.56e-06
|
||||||||
| degP_htrA_DO | TIGR02037 | periplasmic serine protease, Do/DeqQ family; This family consists of a set proteins various ... |
277-502 | 6.67e-06 | ||||
periplasmic serine protease, Do/DeqQ family; This family consists of a set proteins various designated DegP, heat shock protein HtrA, and protease DO. The ortholog in Pseudomonas aeruginosa is designated MucD and is found in an operon that controls mucoid phenotype. This family also includes the DegQ (HhoA) paralog in E. coli which can rescue a DegP mutant, but not the smaller DegS paralog, which cannot. Members of this family are located in the periplasm and have separable functions as both protease and chaperone. Members have a trypsin domain and two copies of a PDZ domain. This protein protects bacteria from thermal and other stresses and may be important for the survival of bacterial pathogens.// The chaperone function is dominant at low temperatures, whereas the proteolytic activity is turned on at elevated temperatures. [Protein fate, Protein folding and stabilization, Protein fate, Degradation of proteins, peptides, and glycopeptides] Pssm-ID: 273938 [Multi-domain] Cd Length: 428 Bit Score: 49.91 E-value: 6.67e-06
|
||||||||
| PDZ4_LNX1_2-like | cd06680 | PDZ domain 4 of human Ligand of Numb protein X 1 (LNX1) and LNX2, and related domains; PDZ ... |
447-494 | 7.24e-06 | ||||
PDZ domain 4 of human Ligand of Numb protein X 1 (LNX1) and LNX2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 4 of LNX1 (also known as PDZ domain-containing RING finger protein 2, PDZRN2)and LNX2 (also known as PDZ domain-containing RING finger protein 1, PDZRN1), and related domains. LNX1 and LNX2 are Ring (Really Interesting New Gene) finger and PDZ domain-containing E3 ubiquitin ligases that bind to the cell fate determinant protein NUMB and mediate its ubiquitination. LNX1 can ubiquitinate a number of other ligands including PPFIA1, KLHL11, KIF7 and ERC2. LNX1 and LNX2 each have four PDZ domains. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This LNX family PDZ4 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467168 [Multi-domain] Cd Length: 89 Bit Score: 45.42 E-value: 7.24e-06
|
||||||||
| PDZ2_GRIP1-2-like | cd06681 | PDZ domain 2 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related ... |
449-488 | 7.69e-06 | ||||
PDZ domain 2 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR) binding proteins GRIP1 (ABP/GRIP2) and GRIP2, and related domains. GRIP1 and GRIP2 each have 7 PDZ domains. The interaction of GRIP1 and GRIP2 with GluA2/3 (AMPAR subunit) regulates AMPAR trafficking and synaptic targeting. GRIP1 has an essential role in regulating AMPAR trafficking during synaptic plasticity and learning and memory. GRIP1 and GRIP2 interact with a variety of other proteins associated with protein trafficking and internalization, for example GRIP1 also interacts with KIF5 (also known as kinesin 1), EphB receptors, scaffold protein liprin-alpha, and the rasGEF GRASP-1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This GRIP family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467169 [Multi-domain] Cd Length: 89 Bit Score: 45.30 E-value: 7.69e-06
|
||||||||
| PDZ4_GRIP1-2-like | cd06686 | PDZ domain 4 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related ... |
449-494 | 8.26e-06 | ||||
PDZ domain 4 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR) binding proteins GRIP1 (ABP/GRIP2) and GRIP2, and related domains. GRIP1 and GRIP2 each have 7 PDZ domains. The interaction of GRIP1 and GRIP2 with GluA2/3 (AMPAR subunit) regulates AMPAR trafficking and synaptic targeting. GRIP1 has an essential role in regulating AMPAR trafficking during synaptic plasticity and learning and memory. GRIP1 and GRIP2 interact with a variety of other proteins associated with protein trafficking and internalization, for example GRIP1 also interacts with KIF5 (also known as kinesin 1), EphB receptors, scaffold protein liprin-alpha, and the rasGEF GRASP-1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This GRIP family PDZ4 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467174 [Multi-domain] Cd Length: 99 Bit Score: 45.41 E-value: 8.26e-06
|
||||||||
| PDZ3_PTPN13_FRMPD2-like | cd06695 | PDZ domain 3 of protein tyrosine phosphatase non-receptor type 13 (PTPN13), FERM and PDZ ... |
252-337 | 8.28e-06 | ||||
PDZ domain 3 of protein tyrosine phosphatase non-receptor type 13 (PTPN13), FERM and PDZ domain-containing protein 2 (FRMPD2), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 3 of PTPN13 [also known as Fas-associated protein-tyrosine phosphatase 1 (FAP-1), protein-tyrosine phosphatase 1E (PTP-E1), and protein-tyrosine phosphatase (PTPL1)], FRMPD2 (also known as PDZ domain-containing protein 4; PDZ domain-containing protein 5C), and related domains. PTPN13 regulates negative apoptotic signaling and mediates phosphoinositide 3-kinase (PI3K) signaling. PTPN13 has five PDZ domains. Proteins known to interact with PTPN13 PDZ domains include: PLEKHA1 and PLEKHA2 via PTPN13-PDZ domain 1, Fas receptor and thyroid receptor-interacting protein 6 via PTPN13-PDZ domain 2, nerve growth factor receptor and protein kinase N2 via PTPN13-PDZ domain 3, PDZ and LIM domain 4 (PDLIM4) via PTPN13-PDZ domains 2 and 4, and brain calpain-2 via PTPN13-PDZ domains 3, 4 and 5. Calpain-2-mediated PTPN13 fragments may be involved in abnormal tau aggregation and increased risk for Alzheimer's disease. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). FRMPD2 is localized in the basolateral membranes of polarized epithelial cells and is associated with tight junction formation and immune response; it contains 3 PDZ domains). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PTPN13 family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467181 [Multi-domain] Cd Length: 90 Bit Score: 45.33 E-value: 8.28e-06
|
||||||||
| PDZ4_MAGI-1_3-like | cd06734 | PDZ domain 4 of membrane-associated guanylate kinase inverted 1 (MAGI-1), MAGI-2, and MAGI-3, ... |
251-327 | 8.76e-06 | ||||
PDZ domain 4 of membrane-associated guanylate kinase inverted 1 (MAGI-1), MAGI-2, and MAGI-3, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 4 of MAGI1, 2, 3 (MAGI is also known as Membrane-associated guanylate kinase, WW and PDZ domain-containing protein) and related domains. MAGI proteins have been implicated in the control of cell migration and invasion through altering the activity of phosphatase and tensin homolog (PTEN) and modulating Akt signaling. Four MAGI proteins have been identified (MAGI1-3 and MAGIX). MAGI1-3 have 6 PDZ domains and bind to the C-terminus of PTEN via their PDZ2 domain. MAGIX has a single PDZ domain that is related to MAGI1-3 PDZ domain 5. Other binding partners for MAGI1 include JAM4, C-terminal tail of high risk HPV-18 E6, megalin, TRAF6, Kir4.1 (basolateral K+ channel subunit), and cadherin 23; for MAGI2, include DASM1, dendrin, axin, beta- and delta-catenin, neuroligin, hyperpolarization-activated cation channels, beta1-adrenergic receptors, NMDA receptor, and TARPs; and for MAGI3 includes LPA2. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MAGI family PDZ4 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as beta-strands A, -B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467216 [Multi-domain] Cd Length: 84 Bit Score: 44.91 E-value: 8.76e-06
|
||||||||
| RING-H2_RNF126-like | cd16667 | RING finger, H2 subclass, found in RING finger proteins RNF126, RNF115, and similar proteins; ... |
18-57 | 9.86e-06 | ||||
RING finger, H2 subclass, found in RING finger proteins RNF126, RNF115, and similar proteins; This subfamily includes RING finger proteins RNF126, RNF115, and similar proteins. RNF126 is a Bag6-dependent E3 ubiquitin ligase that is involved in the mislocalized protein (MLP) pathway of quality control. It regulates the retrograde sorting of the cation-independent mannose 6-phosphate receptor (CI-MPR). RNF126 promotes cancer cell proliferation by targeting the tumor suppressor p21 for ubiquitin-mediated degradation, and could be a novel therapeutic target in breast and prostate cancers. It is also able to ubiquitylate cytidine deaminase (AID), a poorly soluble protein that is essential for antibody diversification. RNF115, also known as Rab7-interacting ring finger protein (Rabring 7), or zinc finger protein 364 (ZNF364), or breast cancer-associated gene 2 (BCA2), is an E3 ubiquitin-protein ligase that is an endogenous inhibitor of adenosine monophosphate-activated protein kinase (AMPK) activation; this inhibition increases the efficacy of metformin in breast cancer cells. It also functions as a cofactor in the restriction imposed by tetherin on HIV-1, and targets HIV-1 Gag for lysosomal degradation, impairing virus assembly and release, in a tetherin-independent manner. Moreover, RNF115 is a Rab7-binding protein that stimulates c-Myc degradation through mono-ubiquitination of MM-1. It also plays crucial roles as a Rab7 target protein in vesicle traffic to late endosome/lysosome and lysosome biogenesis. RNF115 and RNF126 associate with the epidermal growth factor receptor (EGFR) and promote ubiquitylation of EGFR, suggesting they play a role in the ubiquitin-dependent sorting and downregulation of membrane receptors. Both of them contain an N-terminal BCA2 Zinc-finger domain (BZF), AKT-phosphorylation sites, and a C-terminal C3H2C3-type RING-H2 finger. Pssm-ID: 438329 [Multi-domain] Cd Length: 43 Bit Score: 43.45 E-value: 9.86e-06
|
||||||||
| RING-HC_CHFR | cd16503 | RING finger, HC subclass, found in checkpoint with forkhead and RING finger domains protein ... |
14-67 | 1.00e-05 | ||||
RING finger, HC subclass, found in checkpoint with forkhead and RING finger domains protein (CHFR); CHFR, also known as RING finger protein 196 (RNF196), is a checkpoint protein that delays entry into mitosis in response to stress. It functions as an E3 ubiquitin ligase that ubiquitinates and degrades its target proteins, such as Aurora-A, Plk1, Kif22, and PARP-1, which are critical for proper mitotic transitions. It also plays an important role in cell cycle progression and tumor suppression, and is negatively regulated by SUMOylation-mediated proteasomal ubiquitylation. Moreover, CHFR is involved in the early stage of the DNA damage response, which mediates the crosstalk between ubiquitination and poly-ADP-ribosylation. CHFR contains a fork head associated (FHA) domain and a C3HC4-type RING-HC finger. Pssm-ID: 438166 [Multi-domain] Cd Length: 55 Bit Score: 43.89 E-value: 1.00e-05
|
||||||||
| PDZ_SNX27-like | cd23070 | PDZ domain of sorting nexin-27 (SNX27), and related domains; PDZ (PSD-95 (Postsynaptic density ... |
283-336 | 1.07e-05 | ||||
PDZ domain of sorting nexin-27 (SNX27), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of SNX27, and related domains. SNX27 is involved in retrograde transport from endosome to plasma membrane. The PDZ domain of SNX27 links cargo identification to retromer-mediated transport. SNX27 binds to the retromer complex (vacuolar protein sorting 26(VPS26)-VPS29-VPS35), via its PDZ domain binding to VPS26. The SNX27 PDZ domain also binds to cargo including the G-protein-coupled receptors (GPCRs): beta2-adrenergic receptor (beta2AR), beta1AR, parathyroid hormone receptor (PTHR), alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors (AMPARs), NMDA receptors, 5-hydroxytryptamine 4a receptors, frizzled receptors, and somatostatin receptor subtype 5 (SSTR5). Additional binding partners of the SNX27 PDZ domain include G protein-gated inwardly rectifying potassium (Kir3) channels, angiotensin-converting enzyme 2 (ACE2), and PTEN (phosphatase and tensin homolog deleted on chromosome 10); PTEN binding to SNX27 prevents SNX27's association with the retromer complex. SNX27 has been reported to be a host factor needed for efficient entry of an engineered SARS-CoV-2 variant, the spike protein of which contains a deletion at the S1/S2 subunit cleavage site; the PDZ domain of SNX27 binds angiotensin-converting enzyme 2 (ACE2), and may be involved in recycling ACE2 to the plasma membrane, thereby promoting viral entry. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This SNX27-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467283 [Multi-domain] Cd Length: 93 Bit Score: 45.09 E-value: 1.07e-05
|
||||||||
| RING-HC_COP1 | cd16504 | RING finger, HC subclass, found in constitutive photomorphogenesis protein 1 (COP1) and ... |
15-54 | 1.10e-05 | ||||
RING finger, HC subclass, found in constitutive photomorphogenesis protein 1 (COP1) and similar proteins; COP1, also known as RING finger and WD repeat domain protein 2 (RFWD2) or RING finger protein 200 (RNF200), is a central regulator of photomorphogenic development in plants, which targets key transcription factors for proteasome-dependent degradation. It is localized predominantly in the nucleus, but may also be present in the cytosol. Mammalian COP1 functions as an E3 ubiquitin-protein ligase that interacts with Jun transcription factors and modulates their transcriptional activity. It also interacts with and negatively regulates the tumor-suppressor protein p53. Moreover, COP1 associates with COP9 signalosome subunit 6 (CSN6), and is involved in 14-3-3sigma ubiquitin-mediated degradation. The CSN6-COP1 link enhances ubiquitin-mediated degradation of p27(Kip1), a critical CDK inhibitor involved in cell cycle regulation, to promote cancer cell growth. Furthermore, COP1 functions as the negative regulator of ETV1 and influences prognosis in triple-negative breast cancer. COP1 contains an N-terminal extension, a C3HC4-type RING-HC finger, a coiled coil domain, and seven WD40 repeats. In human COP1, a classic leucine-rich NES, and a novel bipartite NLS is bridged by the RING-HC finger. Pssm-ID: 438167 [Multi-domain] Cd Length: 47 Bit Score: 43.38 E-value: 1.10e-05
|
||||||||
| PDZ1_FRMPD2-like | cd23071 | PDZ domain 1 of FERM and PDZ domain-containing protein 2 (FRMPD2), and related domains; PDZ ... |
252-331 | 1.16e-05 | ||||
PDZ domain 1 of FERM and PDZ domain-containing protein 2 (FRMPD2), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of FRMPD2 (also known as PDZ domain-containing protein 4, and related domains. FRMPD2 is localized in the basolateral membranes of polarized epithelial cells and is associated with tight junction formation and immune response; it contains 3 PDZ domains. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PTPN13 family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467284 [Multi-domain] Cd Length: 92 Bit Score: 44.79 E-value: 1.16e-05
|
||||||||
| RING-HC_RNFT2 | cd16742 | RING finger, HC subclass, found in RING finger and transmembrane domain-containing protein 2 ... |
10-57 | 1.22e-05 | ||||
RING finger, HC subclass, found in RING finger and transmembrane domain-containing protein 2(RNFT2); RNFT2, also known as transmembrane protein 118 (TMEM118), is a multi-pass membrane protein containing a C3HC4-type RING-HC finger. Its biological role remains unclear. Pssm-ID: 438400 [Multi-domain] Cd Length: 67 Bit Score: 44.10 E-value: 1.22e-05
|
||||||||
| PDZ1_Dlg1-2-4-like | cd06723 | PDZ domain 1 of human discs large homolog 1 (Dlg1), Dlg2, and Dlg4, Drosophila disc large (Dlg) ... |
431-486 | 1.26e-05 | ||||
PDZ domain 1 of human discs large homolog 1 (Dlg1), Dlg2, and Dlg4, Drosophila disc large (Dlg), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of Drosophila Dlg1, human Dlg1,2, and 4 and related domains. Dlg1 (also known as synapse-associated protein Dlg197 or SAP-97), Dlg2 (also known as channel-associated protein of synapse-110, postsynaptic density protein 93, or PSD-93), Dlg4 (also known as postsynaptic density protein 95, PSD-95, synapse-associated protein 90, or SAP-90) each have 3 PDZ domains and belong to the membrane-associated guanylate kinase family. Dlg1 regulates antigen receptor signaling and cell polarity in lymphocytes, B-cell proliferation and antibody production, and TGFalpha bioavailability; its PDZ3 domain binds pro-TGFalpha, and its PDZ2 domain binds the TACE metalloprotease responsible for cleaving pro-TGFalpha to a soluble form. Dlg2 is involved in N-methyl-D-aspartate (NMDA) receptor signaling. It regulates surface expression of NMDA receptors in dorsal horn neurons of the spinal cord, and it also interacts with NMDA receptor subunits and with Shaker-type K+ channel subunits to cluster into a channel complex. Dlg4 PDZ1 domain binds NMDA receptors, and its PDZ2 domain binds neuronal nitric oxide synthase (nNOS), forming a complex in neurons. The Drosophila Scribble complex (Scribble, Dlg, and lethal giant larvae) plays a role in apico-basal cell polarity, and in other forms of polarity, including regulation of the actin cytoskeleton, cell signaling and vesicular trafficking, and in tumor development. Postsynaptic targeting of Drosophila DLG requires interactions mediated by the first two PDZ domains. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Dlg-like family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467206 [Multi-domain] Cd Length: 89 Bit Score: 44.61 E-value: 1.26e-05
|
||||||||
| PDZ_Par6-like | cd06718 | PDZ domain of partitioning defective 6 (Par6), Drosophila Rho GTPase-activating protein 100F ... |
420-480 | 1.34e-05 | ||||
PDZ domain of partitioning defective 6 (Par6), Drosophila Rho GTPase-activating protein 100F (RhoGAP100F), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of Par6 (also known as PAR6 or Par-6), RhoGAP100F, and related domains. Par6 is part of a conserved machinery that directs metazoan cell polarity, a process necessary for the function of diverse cell types. Par6 forms a cell polarity-regulatory complex with atypical protein kinase C (aPKC) and Par3. Par6 can also directly associate with PALS1 (proteins associated with Lin7, also known as Stardust) providing a link between the Par3/aPKC/Par6 complex and the PALS1-PATJ (protein-associated TJ) complex. Binding partners of the Par6-PDZ domain include Par3, PALS1/Stardust; leucine-rich repeat-containing protein netrin-G ligand-2 (NGL-2), human crumbs (CRB3) involve in the morphogenesis of the tight junctions in mammalian epithelial cells, and PAR-6 co-operates with the Par6 semi-CRIB domain to bind CDC42. CDC42 regulates the Par6 PDZ domain through an allosteric CRIB-PDZ transition. Drosophila RhoGAP100F, also known as synapse defective protein 1 homolog (syd-1 homolog), is a GTPase activator for the Rho-type GTPases by converting them to an inactive GDP-bound form. The RhoGAP100F-PDZ domain binds the neurexin C terminus to control synapse formation at the Drosophila neuromuscular junction. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Par6-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467202 [Multi-domain] Cd Length: 84 Bit Score: 44.48 E-value: 1.34e-05
|
||||||||
| PDZ_rhophilin-like | cd06712 | PDZ domain of rhophilin-1, rhophilin-2, and related domains; PDZ (PSD-95 (Postsynaptic density ... |
249-335 | 1.58e-05 | ||||
PDZ domain of rhophilin-1, rhophilin-2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of rhophilin-1, rhophilin-2, and related domains. Rhophilin-1 (RHPN1, also known as GTP-Rho-binding protein 1) and rhophilin-2 (RHPN2, also known as GTP-Rho-binding protein 2) are Rho-GTP binding proteins involved in cytoskeletal dynamics. Rhophilin-2 inhibits RhoA's activity to induce F-actin stress fibers. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This rhophilin-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467196 [Multi-domain] Cd Length: 78 Bit Score: 44.11 E-value: 1.58e-05
|
||||||||
| PDZ2_PDZD7-like | cd10834 | PDZ domain 2 of the canonical isoform 1 of PDZ domain containing 7 (PDZD7), and related ... |
420-491 | 1.68e-05 | ||||
PDZ domain 2 of the canonical isoform 1 of PDZ domain containing 7 (PDZD7), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of the long isoform 1 of PDZD7, and related domains. PDZD7 is critical for the organization of Usher syndrome type 2 (USH2) complex. Usher syndrome is the leading cause of hereditary sensory deaf-blindness in humans; USH2 is the most common sub-type. Formation of the USH2 complex is based upon heterodimerization between PDZD7 and whirlin (another PDZ domain-containing protein) and a subsequent dynamic interplay between USH2 proteins via their multiple PDZ domains. The PDZD7 PDZ2 domain binds GPR98 (also known as VLGR1) and usherin (USH2A). PDZD7 and whirlin form heterodimers through their multiple PDZ domains; whirlin and PDZD7 interact with usherin and GPR98 to form an interdependent ankle link complex. PDZD7 also interacts with myosin VIIa. PDZD7 also forms homodimers through its PDZ2 domain. Various isoforms of PDZD7 produced by alternative splicing have been identified; this subgroup includes the second PDZ domain of the canonical isoform of PDZD7- isoform 1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZD7-like family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467270 [Multi-domain] Cd Length: 85 Bit Score: 44.30 E-value: 1.68e-05
|
||||||||
| PDZ_Dishevelled-like | cd06717 | PDZ domain of segment polarity protein dishevelled homolog DVL1, DVL2, DVL3, and related ... |
259-335 | 1.69e-05 | ||||
PDZ domain of segment polarity protein dishevelled homolog DVL1, DVL2, DVL3, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of DVL1-3, and related domains. The dishevelleds (DVL1, 2 and 3 in humans) act downstream of Frizzled (FZD) receptors in both the canonical and non-canonical WNT signaling pathway; they bind the cytoplasmic C-terminus of frizzled family members and transduce the Wnt signal to down-stream effectors. They bind to several proteins known to modulate Wnt signaling. Binding partners of the DVL1 PDZ domain include nucleoredoxin (NXN), Van Gogh-like (VANGL1), Wnt receptor RYK, Dapper 1 (DACT1), Frizzled7 (FZD7), transmembrane protein 88 (TMEM88), Daple (dishevelled-associating protein with a high frequency of leucines), also known as Ccdc88c), and cysteine-rich protein Idax. The DVL2 PDZ domain has been shown to bind the nuclear export signal sequence of the DVL2 protein. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This DVL-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467201 [Multi-domain] Cd Length: 87 Bit Score: 44.28 E-value: 1.69e-05
|
||||||||
| PDZ1_FRMPD2-like | cd23071 | PDZ domain 1 of FERM and PDZ domain-containing protein 2 (FRMPD2), and related domains; PDZ ... |
444-492 | 1.75e-05 | ||||
PDZ domain 1 of FERM and PDZ domain-containing protein 2 (FRMPD2), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of FRMPD2 (also known as PDZ domain-containing protein 4, and related domains. FRMPD2 is localized in the basolateral membranes of polarized epithelial cells and is associated with tight junction formation and immune response; it contains 3 PDZ domains. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PTPN13 family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467284 [Multi-domain] Cd Length: 92 Bit Score: 44.41 E-value: 1.75e-05
|
||||||||
| zf-RING_UBOX | pfam13445 | RING-type zinc-finger; This zinc-finger is a typical RING-type of plant ubiquitin ligases. |
18-53 | 1.81e-05 | ||||
RING-type zinc-finger; This zinc-finger is a typical RING-type of plant ubiquitin ligases. Pssm-ID: 463881 [Multi-domain] Cd Length: 38 Bit Score: 42.77 E-value: 1.81e-05
|
||||||||
| PDZ_densin_erbin-like | cd06749 | PDZ domain of densin, erbin, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95) ... |
443-502 | 1.84e-05 | ||||
PDZ domain of densin, erbin, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of densin, erbin, and related domains. Densin (also known as leucine-rich repeat-containing protein 7, LRRC7, densin-180, protein LAP1) and erbin (also known as densin-180-like protein, Erbb2-interacting protein, protein LAP2) belong to the LAP (leucine-rich repeat and PDZ domain) family of scaffolding proteins that play roles in the maintenance of cell shape and apical-basal polarity. Densin and erbin are components of the excitatory postsynaptic compartment and are regulators of dendritic morphology and postsynaptic structure. The densin PDZ domain binds CaV1.3 alpha1 subunit, delta-catenin, and MAGUIN-1. Binding partners of the erbin PDZ domain include ErbB receptor tyrosine kinase ErbB2, HTLV-1 Tax1, Cav1.3 Ca2+channels, and constituents of the cadherin:catenin cell adhesion complex, in particular delta-catenin, p0071 and ARVCF. The erbin PDZ domain binds Smad3, a transductor of the TGFbeta pathway, possibly by a novel interface of binding. Erbin and two other LAP proteins (scribble and lano) redundantly regulate epithelial polarity and apical adhesion complex. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This densin and erbin-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged as beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467231 [Multi-domain] Cd Length: 87 Bit Score: 44.24 E-value: 1.84e-05
|
||||||||
| PDZ3_MAGI-1_3-like | cd06733 | PDZ domain 3 of membrane-associated guanylate kinase inverted 1 (MAGI-1), MAGI-2, and MAGI-3, ... |
441-490 | 1.97e-05 | ||||
PDZ domain 3 of membrane-associated guanylate kinase inverted 1 (MAGI-1), MAGI-2, and MAGI-3, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 3 of MAGI1, 2, 3 (MAGI is also known as Membrane-associated guanylate kinase, WW and PDZ domain-containing protein) and related domains. MAGI proteins have been implicated in the control of cell migration and invasion through altering the activity of phosphatase and tensin homolog (PTEN) and modulating Akt signaling. Four MAGI proteins have been identified (MAGI1-3 and MAGIX). MAGI1-3 have 6 PDZ domains and bind to the C-terminus of PTEN via their PDZ2 domain. MAGIX has a single PDZ domain that is related to MAGI1-3 PDZ domain 5. Other binding partners for MAGI1 include JAM4, C-terminal tail of high risk HPV-18 E6, megalin, TRAF6, Kir4.1 (basolateral K+ channel subunit), and cadherin 23; for MAGI2, include DASM1, dendrin, axin, beta- and delta-catenin, neuroligin, hyperpolarization-activated cation channels, beta1-adrenergic receptors, NMDA receptor, and TARPs; and for MAGI3 includes LPA2. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MAGI family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as beta-strands A, -B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467215 [Multi-domain] Cd Length: 85 Bit Score: 43.75 E-value: 1.97e-05
|
||||||||
| RING-HC_TRIM72_C-IV | cd16612 | RING finger, HC subclass, found in tripartite motif-containing protein 72 (TRIM72) and similar ... |
15-41 | 2.17e-05 | ||||
RING finger, HC subclass, found in tripartite motif-containing protein 72 (TRIM72) and similar proteins; TRIM72, also known as Mitsugumin-53 (MG53), is a muscle-specific protein that plays a central role in cell membrane repair by nucleating the assembly of the repair machinery at muscle injury sites. It is required in repair of alveolar epithelial cells under plasma membrane stress failure. It interacts with dysferlin to regulate sarcolemmal repair. Upregulation of TRIM72 develops obesity, systemic insulin resistance, dyslipidemia, and hyperglycemia, as well as induces diabetic cardiomyopathy through transcriptional activation of the peroxisome proliferation-activated receptor alpha (PPAR-alpha) signaling pathway. Compensation for the absence of AKT signaling by ERK signaling during TRIM72 overexpression leads to pathological hypertrophy. Moreover, TRIM72 functions as a novel negative feedback regulator of myogenesis by targeting insulin receptor substrate-1 (IRS-1). It is transcriptionally activated by the synergism of myogenin (MyoD) and myocyte enhancer factor 2 (MEF2). TRIM72 belongs to the C-IV subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region, as well as a B30.2/SPRY (SplA and ryanodine receptor) domain positioned C-terminal to the RBCC domain. Pssm-ID: 438274 [Multi-domain] Cd Length: 60 Bit Score: 43.19 E-value: 2.17e-05
|
||||||||
| mRING-HC-C3HC3D_TRAF7 | cd16644 | Modified RING finger, HC subclass (C3HC3D-type), found in tumor necrosis factor (TNF) ... |
16-53 | 2.30e-05 | ||||
Modified RING finger, HC subclass (C3HC3D-type), found in tumor necrosis factor (TNF) receptor-associated factor 7 (TRAF7) and similar proteins; TRAF7, also known as RING finger and WD repeat-containing protein 1 or RING finger protein 119 (RNF119), is an E3 ubiquitin-protein ligase involved in signal transduction pathways that lead either to activation or repression of NF-kappaB transcription factor by promoting K29-linked ubiquitination of several cellular targets, including the NF-kappaB essential modulator (NEMO) and the p65 subunit of NF-kappaB transcription factor. It is also involved in K29-linked polyubiquitination that has been implicated in lysosomal degradation of proteins. Moreover, TRAF7 is required for K48-linked ubiquitination of p53, a key tumor suppressor and a master regulator of various signaling pathways, such as those related to apoptosis, cell cycle and DNA repair. It is also required for tumor necrosis factor alpha (TNFalpha)-induced Jun N-terminal kinase activation and promotes cell death by regulating polyubiquitination and lysosomal degradation of c-FLIP protein. Furthermore, TRAF7 functions as small ubiquitin-like modifier (SUMO) E3 ligase involved in other post-translational modification, such as sumoylation. It binds to and stimulates sumoylation of the proto-oncogene product c-Myb, a transcription factor regulating proliferation and differentiation of hematopoietic cells. It potentiates MEKK3-induced AP1 and CHOP activation and induces apoptosis. Meanwhile, TRAF7 mediates MyoD1 regulation of the pathway and cell-cycle progression in myoblasts. It also plays a role in Toll-like receptors (TLR) signaling. TRAF7 contains an N-terminal domain with a modified C3HC3D-type RING-HC finger and an adjacent zinc finger, and a unique C-terminal domain that comprises a coiled coil domain and seven WD40 repeats. Pssm-ID: 438306 [Multi-domain] Cd Length: 47 Bit Score: 42.73 E-value: 2.30e-05
|
||||||||
| PDZ_RGS12-like | cd06710 | PDZ domain of regulator of G-protein signaling 12 (RGS12), and related domains; PDZ (PSD-95 ... |
259-321 | 2.42e-05 | ||||
PDZ domain of regulator of G-protein signaling 12 (RGS12), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of RGS12, and related domains. RGS12 downregulates GPCR signal transduction by increasing the GTPase activity of G-protein alpha subunits, thereby driving G-proteins into their inactive GDP-bound form. The RGS12 PDZ domain can bind selectively to C-terminal (A/S)-T-X-(L/V) motifs as found within both the CXCR2 IL-8 receptor, and the alternative 3' exon form of RGS12. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This RGS12-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467194 [Multi-domain] Cd Length: 76 Bit Score: 43.39 E-value: 2.42e-05
|
||||||||
| RING-HC_EHV1-like | cd23130 | RING finger, HC subclass, found in Equid alphaherpesvirus 1 (Equine herpesvirus 1/EHV-1) ... |
18-61 | 2.49e-05 | ||||
RING finger, HC subclass, found in Equid alphaherpesvirus 1 (Equine herpesvirus 1/EHV-1) regulatory protein and similar proteins; EHV-1 regulatory protein belongs to the Vmw110 (IPC0) protein family. It contains a typical C3HC4-type RING-HC finger and binds zinc stably. Pssm-ID: 438492 [Multi-domain] Cd Length: 51 Bit Score: 42.73 E-value: 2.49e-05
|
||||||||
| RING-HC_TRIM7-like_C-IV | cd16594 | RING finger, HC subclass, found in tripartite motif-containing proteins, TRIM7, TRIM11 and ... |
15-57 | 2.74e-05 | ||||
RING finger, HC subclass, found in tripartite motif-containing proteins, TRIM7, TRIM11 and TRIM27, and similar proteins; TRIM7, TRIM11 and TRIM27, closely related tripartite motif-containing proteins, belong to the C-IV subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox2, and a coiled coil region, as well as a SPRY/B30.2 domain positioned C-terminal to the RBCC domain. TRIM7, also known as glycogenin-interacting protein (GNIP) or RING finger protein 90 (RNF90), is an E3 ubiquitin-protein ligase that mediates c-Jun/AP-1 activation by Ras signalling. Its phosphorylation and activation by MSK1 in response to direct activation by the Ras-Raf-MEK-ERK pathway can stimulate TRIM7 E3 ubiquitin ligase activity in mediating Lys63-linked ubiquitination of the AP-1 coactivator RACO-1, leading to RACO-1 protein stabilization. Moreover, TRIM7 binds and activates glycogenin, the self-glucosylating initiator of glycogen biosynthesis. TRIM11, also known as protein BIA1, or RING finger protein 92 (RNF92), is an E3 ubiquitin-protein ligase involved in the development of the central nervous system. It is overexpressed in high-grade gliomas and promotes proliferation, invasion, migration and glial tumor growth. TRIM11 acts as a potential therapeutic target for congenital central hypoventilation syndrome (CCHS) by mediating the degradation of CCHS-associated polyalanine-expanded Phox2b. TRIM11 modulates the function of neurogenic transcription factor Pax6 through the ubiquitin-proteosome system, and thus plays an essential role for Pax6-dependent neurogenesis. It also binds to and destabilizes a key component of the activator-mediated cofactor complex (ARC105), humanin, a neuroprotective peptide against Alzheimer's disease-relevant insults, and further regulates ARC105 function in transforming growth factor beta (TGFbeta) signaling. Moreover, TRIM11 negatively regulates retinoic acid-inducible gene-I (RIG-I)-mediated interferon-beta (IFNbeta) production and antiviral activity by targeting TANK-binding kinase-1 (TBK1). It may contribute to the endogenous restriction of retroviruses in cells. It enhances N-tropic murine leukemia virus (N-MLV) entry by interfering with Ref1 restriction. It also suppresses the early steps of human immunodeficiency virus HIV-1 transduction, resulting in decreased reverse transcripts. TRIM27, also known as RING finger protein 76 (RNF76), RET finger protein (RFP), or zinc finger protein RFP, is a nuclear E3 ubiquitin-protein ligase that is highly expressed in testis and in various tumor cell lines. Expression of TRIM27 is associated with prognosis of colon and endometrial cancers. TRIM27 was first identified as a fusion partner of the RET receptor tyrosine kinase. It functions as a transcriptional repressor and associates with several proteins involved in transcriptional activity, such as enhancer of polycomb 1 (Epc1), a member of the Polycomb group proteins, and Mi-2beta, a main component of the nucleosome remodeling and deacetylase (NuRD) complex, and the cell cycle regulator retinoblastoma protein (RB1). It also interacts with HDAC1, leading to downregulation of thioredoxin binding protein 2 (TBP-2), which inhibits the function of thioredoxin. Moreover, TRIM27 mediates Pax7-induced ubiquitination of MyoD in skeletal muscle atrophy. In addition, it inhibits muscle differentiation by modulating serum response factor (SRF) and Epc1. TRIM27 promotes a non-canonical polyubiquitination of PTEN, a lipid phosphatase that catalyzes PtdIns(3,4,5)P3 (PIP3) to PtdIns(4,5)P2 (PIP2). It is an IKKepsilon-interacting protein that regulates IkappaB kinase (IKK) function and negatively regulates signaling involved in the antiviral response and inflammation. TRIM27 also forms a protein complex with MBD4 or MBD2 or MBD3, and thus plays an important role in the enhancement of transcriptional repression through MBD proteins in tumorigenesis, spermatogenesis, and embryogenesis. It is a component of an estrogen receptor 1 (ESR1) regulatory complex that is involved in estrogen receptor-mediated transcription in MCF-7 cells. Pssm-ID: 438256 [Multi-domain] Cd Length: 61 Bit Score: 42.67 E-value: 2.74e-05
|
||||||||
| PDZ7_GRIP1-2-like | cd06685 | PDZ domain 7 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related ... |
248-312 | 2.74e-05 | ||||
PDZ domain 7 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR) binding proteins GRIP1 (ABP/GRIP2) and GRIP2, and related domains. GRIP1 and GRIP2 each have 7 PDZ domains. The interaction of GRIP1 and GRIP2 with GluA2/3 (AMPAR subunit) regulates AMPAR trafficking and synaptic targeting. GRIP1 has an essential role in regulating AMPAR trafficking during synaptic plasticity and learning and memory. GRIP1 and GRIP2 interact with a variety of other proteins associated with protein trafficking and internalization, for example GRIP1 also interacts with KIF5 (also known as kinesin 1), EphB receptors, scaffold protein liprin-alpha, and the rasGEF GRASP-1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This GRIP family PDZ7 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467173 [Multi-domain] Cd Length: 85 Bit Score: 43.40 E-value: 2.74e-05
|
||||||||
| cpPDZ1_DegP-like | cd10839 | circularly permuted first PDZ domain (PDZ1) of Escherichia coli periplasmic serine ... |
276-337 | 2.76e-05 | ||||
circularly permuted first PDZ domain (PDZ1) of Escherichia coli periplasmic serine endoprotease DegP and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of Escherichia coli DegP (also known as heat shock protein DegP and Protease Do) and related domains. DegP belongs to the HtrA family of housekeeping proteases. It acts as a protease, degrading transiently denatured and unfolded or misfolded proteins which accumulate in the periplasm following heat shock or other stress conditions, and as a molecular chaperone at low temperatures. DegP has two PDZ domains in addition to the protease domain; its PDZ1 domain is responsible for identifying the distinct substrate sequences that affect degradation (degron) of the substrate sequence, and its PDZ2 domain is responsible for combining with other DegP monomers to form a stable oligomer structure. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains and as well as those with circular permutations and domain swapping of beta-strands. The canonical PDZ domain contains six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. This DegP family PDZ domain 1 is a circularly permuted PDZ domain which places beta-strand A on the C-terminus. Another permutation exists in the PDZ superfamily which places both beta-strands A and B on the C-terminus. Pssm-ID: 467630 [Multi-domain] Cd Length: 91 Bit Score: 43.62 E-value: 2.76e-05
|
||||||||
| RING-HC_RNF168 | cd16550 | RING finger, HC subclass, found in RING finger protein 168 (RNF168) and similar proteins; ... |
18-61 | 2.83e-05 | ||||
RING finger, HC subclass, found in RING finger protein 168 (RNF168) and similar proteins; RNF168 is an E3 ubiquitin-protein ligase that promotes noncanonical K27 ubiquitination to signal DNA damage. It, together with RNF8, functions as a DNA damage response (DDR) factor that promotes a series of ubiquitylation events on substrates, such as H2A and H2AX with H2AK13/15 ubiquitylation, facilitates recruitment of repair factors p53-binding protein 1 (53BP1) or the RAP80-BRCA1 complex to sites of double-strand breaks (DSBs), and inhibits homologous recombination (HR) in cells deficient in the tumor suppressor BRCA1. RNF168 also promotes H2A neddylation, which antagonizes ubiquitylation of H2A and regulates DNA damage repair. Moreover, RNF168 forms a functional complex with RAD6A or RAD6B during the DNA damage response. RNF168 contains an N-terminal C3HC4-type RING-HC finger that catalyzes H2A-K15ub and interacts with H2A, and two MIU (motif interacting with ubiquitin) domains responsible for the interaction with K63 linked poly-ubiquitin. Pssm-ID: 438212 [Multi-domain] Cd Length: 48 Bit Score: 42.36 E-value: 2.83e-05
|
||||||||
| PDZ_tamalin_CYTIP-like | cd06713 | PDZ domain of tamalin, cytohesin-1-interacting protein (CYTIP), and related domains; PDZ ... |
249-327 | 2.92e-05 | ||||
PDZ domain of tamalin, cytohesin-1-interacting protein (CYTIP), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of tamalin, cytohesin-1-interacting protein, and related domains. Tamalin (trafficking regulator and scaffold protein tamalin, also known as general receptor for phosphoinositides 1-associated scaffold protein, GRASP) functions to link receptors, including group 1 metabotropic glutamate receptors (mGluRs), to neuronal proteins. The tamalin PDZ domain binds the C-terminal domains of group I mGluRs; it also binds potassium/sodium hyperpolarization-activated cyclic nucleotide-gated channel 2 (HCN2), neurotrophin-3 (NT3) TrkCT1-truncated receptor, SAP90/PSD-95-associated protein, and tamalin itself. CYTIP (cytohesin-1-interacting protein, also known as Pleckstrin homology Sec7 and coiled-coil domain-binding protein) sequesters cytohesin-1 in the cytoplasm, limiting its interaction with beta2 integrins; cytohesin-1 binds the CYTIP coiled coil domain. The CYTIP PDZ domain can bind the C-terminal peptide of protocadherin alpha-1 (PCDHA1), indicating a possible interaction between the two. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This tamalin-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467197 [Multi-domain] Cd Length: 91 Bit Score: 43.77 E-value: 2.92e-05
|
||||||||
| RseP | COG0750 | Membrane-associated protease RseP, regulator of RpoE activity [Posttranslational modification, ... |
449-502 | 3.09e-05 | ||||
Membrane-associated protease RseP, regulator of RpoE activity [Posttranslational modification, protein turnover, chaperones, Transcription]; Pssm-ID: 440513 [Multi-domain] Cd Length: 349 Bit Score: 47.39 E-value: 3.09e-05
|
||||||||
| PDZ5_PTPN13-like | cd06697 | PDZ domain 5 of protein tyrosine phosphatase non-receptor type 13 (PTPN13), and related ... |
446-502 | 3.14e-05 | ||||
PDZ domain 5 of protein tyrosine phosphatase non-receptor type 13 (PTPN13), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 5 of PTPN13 [also known as Fas-associated protein-tyrosine phosphatase 1 (FAP-1), Protein-tyrosine phosphatase 1E (PTP-E1), and Protein-tyrosine phosphatase (PTPL1)] and related domains. PTPN13 regulates negative apoptotic signaling and mediates phosphoinositide 3-kinase (PI3K) signaling. PTPN13 has five PDZ domains. Proteins known to interact with PTPN13 PDZ domains include: PLEKHA1 and PLEKHA2 via PTPN13-PDZ domain 1, Fas receptor and thyroid receptor-interacting protein 6 via PTPN13-PDZ domain 2, nerve growth factor receptor and protein kinase N2 via PTPN13-PDZ domain 3, PDZ and LIM domain 4 (PDLIM4) via PTPN13-PDZ domains 2 and 4, and brain calpain-2 via PTPN13-PDZ domains 3, 4 and 5. Calpain-2-mediated PTPN13 fragments may be involved in abnormal tau aggregation and increased risk for Alzheimer's disease. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PTPN13 family PDZ5 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467183 [Multi-domain] Cd Length: 87 Bit Score: 43.49 E-value: 3.14e-05
|
||||||||
| RING-HC_RNFT1 | cd16741 | RING finger, HC subclass, found in RING finger and transmembrane domain-containing protein 1 ... |
13-57 | 3.16e-05 | ||||
RING finger, HC subclass, found in RING finger and transmembrane domain-containing protein 1 (RNFT1); RNFT1, also known as protein PTD016, is a multi-pass membrane protein containing a C3HC4-type RING-HC finger. Its biological role remains unclear. Pssm-ID: 438399 [Multi-domain] Cd Length: 58 Bit Score: 42.57 E-value: 3.16e-05
|
||||||||
| RING-H2_RNF181 | cd16669 | RING finger, H2 subclass, found in RING finger protein 181 (RNF181) and similar proteins; ... |
17-60 | 3.19e-05 | ||||
RING finger, H2 subclass, found in RING finger protein 181 (RNF181) and similar proteins; RNF181, also known as HSPC238, is a platelet E3 ubiquitin-protein ligase containing a C3H2C3-type RING-H2 finger. It interacts with the KVGFFKR motif of platelet integrin alpha(IIb)beta3, suggesting a role for RNF181-mediated ubiquitination in integrin and platelet signaling. It also suppresses the tumorigenesis of hepatocellular carcinoma (HCC) through the inhibition of extracellular signal-regulated kinase/mitogen-activated protein kinase (ERK/MAPK) signaling in the liver. Pssm-ID: 438331 [Multi-domain] Cd Length: 46 Bit Score: 41.97 E-value: 3.19e-05
|
||||||||
| PDZ9_MUPP1-like | cd10817 | PDZ domain 9 of multi-PDZ-domain protein 1 (MUPP1) and related domains; PDZ (PSD-95 ... |
428-474 | 3.24e-05 | ||||
PDZ domain 9 of multi-PDZ-domain protein 1 (MUPP1) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 9 of MUPP1. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, PDZ9, and PDZ13. This MuPP1-like PDZ9 domain is therefore absent from PATJ. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ9 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467263 [Multi-domain] Cd Length: 79 Bit Score: 43.11 E-value: 3.24e-05
|
||||||||
| RING-HC_TRIM59_C-V | cd16763 | RING finger, HC subclass, found in tripartite motif-containing protein 59 (TRIM59) and similar ... |
15-42 | 3.39e-05 | ||||
RING finger, HC subclass, found in tripartite motif-containing protein 59 (TRIM59) and similar proteins; TRIM59, also known as RING finger protein 104 (RNF104) or tumor suppressor TSBF-1, is a putative E3 ubiquitin-protein ligase that functions as a novel multiple cancer biomarker for immunohistochemical detection of early tumorigenesis. It is upregulated in gastric cancer and promotes gastric carcinogenesis by interacting with and targeting the P53 tumor suppressor for its ubiquitination and degradation. It also acts as a novel accessory molecule involved in cytotoxicity of BCG-activated macrophages (BAM). Moreover, TRIM59 may serve as a multifunctional regulator for innate immune signaling pathways. It interacts with ECSIT and negatively regulates nuclear factor-kappaB (NF- kappa B) and interferon regulatory factor (IRF)-3/7-mediated signal pathways. TRIM59 belongs to the C-V subclass of the TRIM (tripartite motif) family of proteins that are defined by an N-terminal RBCC (RING, Bbox, and coiled coil) domain, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region. In addition, TRIM59 contains a C-terminal transmembrane domain. Pssm-ID: 438419 [Multi-domain] Cd Length: 56 Bit Score: 42.59 E-value: 3.39e-05
|
||||||||
| PDZ1_PTPN13-like | cd23072 | PDZ domain 1 of protein tyrosine phosphatase non-receptor type 13 (PTPN13), and related ... |
443-506 | 3.57e-05 | ||||
PDZ domain 1 of protein tyrosine phosphatase non-receptor type 13 (PTPN13), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of PTPN13 [also known as Fas-associated protein-tyrosine phosphatase 1 (FAP-1), protein-tyrosine phosphatase 1E (PTP-E1), and protein-tyrosine phosphatase (PTPL1)], and related domains. PTPN13 regulates negative apoptotic signaling and mediates phosphoinositide 3-kinase (PI3K) signaling. PTPN13 has five PDZ domains. Proteins known to interact with PTPN13 PDZ domains include: PLEKHA1 and PLEKHA2 via PTPN13-PDZ domain 1, Fas receptor and thyroid receptor-interacting protein 6 via PTPN13-PDZ domain 2, nerve growth factor receptor and protein kinase N2 via PTPN13-PDZ domain 3, PDZ and LIM domain 4 (PDLIM4) via PTPN13-PDZ domains 2 and 4, and brain calpain-2 via PTPN13-PDZ domains 3, 4 and 5. Calpain-2-mediated PTPN13 fragments may be involved in abnormal tau aggregation and increased risk for Alzheimer's disease. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PTPN13 family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467285 [Multi-domain] Cd Length: 92 Bit Score: 43.63 E-value: 3.57e-05
|
||||||||
| PDZ4_MUPP1-like | cd06668 | PDZ domain 4 of multi-PDZ-domain protein 1 (MUPP1) and PATJ (protein-associated tight junction) ... |
431-488 | 3.63e-05 | ||||
PDZ domain 4 of multi-PDZ-domain protein 1 (MUPP1) and PATJ (protein-associated tight junction) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 4 of MUPP1 and PATJ, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ4 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F Pssm-ID: 467156 [Multi-domain] Cd Length: 88 Bit Score: 43.44 E-value: 3.63e-05
|
||||||||
| PDZ6_PDZD2-PDZ3_hPro-IL-16-like | cd06762 | PDZ domain 6 of PDZ domain containing 2 (PDZD2), PDZ domain 3 of human pro-interleukin-16 ... |
431-502 | 4.11e-05 | ||||
PDZ domain 6 of PDZ domain containing 2 (PDZD2), PDZ domain 3 of human pro-interleukin-16 (isoform 1, 1332 AA), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 6 of PDZD2, also known as KIAA0300, PIN-1, activated in prostate cancer (AIPC) and PDZ domain-containing protein 3 (PDZK3). PDZD2 has seven PDZ domains. PDZD2 is expressed at exceptionally high levels in the pancreas and certain cancer tissues, such as prostate cancer. It promotes the proliferation of insulinoma cells and is upregulated during prostate tumorigenesis. In osteosarcoma (OS), the microRNA miR-363 acts as a tumor suppressor by inhibiting PDZD2. This family also includes the third PDZ domain (PDZ3) of human pro-interleukin-16 (isoform 1, also known as nPro-IL-16). Precursor IL-16 is cleaved to produce pro-IL-16 and C-terminal mature IL-16. Pro-IL-16 functions as a regulator of T cell growth; mature IL-16 is a CD4 ligand that induces chemotaxis and CD25 expression in CD4+ T cells. IL-16 bioactivity has been closely associated with the progression of several different cancers. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZD2-like family PDZ6 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467243 [Multi-domain] Cd Length: 86 Bit Score: 43.02 E-value: 4.11e-05
|
||||||||
| PDZ1_INAD-like | cd23063 | PDZ domain 1 of inactivation-no-after-potential D (INAD), and related domains; PDZ (PSD-95 ... |
446-487 | 4.51e-05 | ||||
PDZ domain 1 of inactivation-no-after-potential D (INAD), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of INAD, and related domains. INAD assembles key enzymes of the Drosophila compound eye photo-transduction pathway into a supramolecular complex, supporting efficient and fast light signaling. It contains 5 PDZ domains arranged in tandem (PDZ1-PDZ5) which independently bind various proteins. INAD PDZ2 binds eye-specific protein kinase C, INAD PDZ3 binds transient receptor potential (TRP) channel, and INAD PDZ4,5 tandem binds NORPA (phospholipase Cbeta, PLCbeta). Mutations of the inaD gene that lead to disruption of each of these interactions impair fly photo signal transduction. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This INAD-like family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467276 [Multi-domain] Cd Length: 87 Bit Score: 42.89 E-value: 4.51e-05
|
||||||||
| PDZ10_MUPP1-PDZ8_PATJ-like | cd06673 | PDZ domain 10 of multi-PDZ-domain protein 1 (MUPP1), domain 8 of PATJ (protein-associated ... |
447-488 | 4.56e-05 | ||||
PDZ domain 10 of multi-PDZ-domain protein 1 (MUPP1), domain 8 of PATJ (protein-associated tight junction) and similar domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 10 of MUPP1, PDZ domain 8 of PATJ, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ10 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467161 [Multi-domain] Cd Length: 86 Bit Score: 43.05 E-value: 4.56e-05
|
||||||||
| RING-HC_ScRAD18-like | cd23148 | RING finger, HC subclass, found in Saccharomyces cerevisiae radiation sensitivity protein 18 ... |
16-65 | 4.62e-05 | ||||
RING finger, HC subclass, found in Saccharomyces cerevisiae radiation sensitivity protein 18 (RAD18) and similar proteins; RAD18, also called RING-type E3 ubiquitin transferase RAD18, acts as a postreplication repair E3 ubiquitin-protein ligase that associates with the E2 ubiquitin conjugating enzyme UBC2/RAD6 to form the UBC2-RAD18 ubiquitin ligase complex involved in postreplicative repair (PRR) of damaged DNA. The UBC2-RAD18 complex cooperates with RAD5 and the UBC13-MMS2 dimer to attach mono-ubiquitin chains on 'Lys-164' of POL30, which is necessary for PRR. The UBC2-RAD18 complex is also involved in prevention of spontaneous mutations caused by 7,8-dihydro-8-oxoguanine. RAD18 is an E3 RING-finger protein belonging to the UBC2/RAD6 epistasis group. It contains a typical C3HC4-type RING-HC finger. Pssm-ID: 438510 [Multi-domain] Cd Length: 52 Bit Score: 41.75 E-value: 4.62e-05
|
||||||||
| RING-HC_RAD16-like | cd16567 | RING finger, HC subclass, found in Saccharomyces cerevisiae DNA repair protein RAD16, ... |
18-54 | 5.33e-05 | ||||
RING finger, HC subclass, found in Saccharomyces cerevisiae DNA repair protein RAD16, Schizosaccharomyces pombe rhp16, and similar proteins; Budding yeast RAD16, also known as ATP-dependent helicase RAD16, is encoded by a yeast excision repair gene homologous to the recombinational repair gene RAD54 and to the SNF2 gene involved in transcriptional activation. It is a component of the global genome repair (GGR) complex that promotes global genome nucleotide excision repair (GG-NER) by removing DNA damage from non-transcribing DNA. RAD16 is involved in differential repair of DNA after UV damage, and repairs preferentially the MAT-alpha locus compared with the HML-alpha locus. Fission yeast rhp16, also known as ATP-dependent helicase rhp16, is a RAD16 homolog. It is involved in GGR via nucleotide excision repair (NER), in conjunction with rhp7, after UV irradiation. Both RAD16 and rhp16 contain a C3HC4-type RING-HC finger, as well as a DEAD-like helicase domain and a helicase superfamily C-terminal domain. Pssm-ID: 438229 [Multi-domain] Cd Length: 48 Bit Score: 41.56 E-value: 5.33e-05
|
||||||||
| PDZ2_harmonin | cd06738 | PDZ domain 2 of harmonin isoforms a, b, and c, and related domains; PDZ (PSD-95 (Postsynaptic ... |
446-490 | 5.64e-05 | ||||
PDZ domain 2 of harmonin isoforms a, b, and c, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of harmonin isoforms a, b, and c, and related domains. Harmonin (also known as Usher Type 1C, PDZ-73 and AIE-75) is a key organizer of the Usher (USH) protein interactome. USH syndrome is the leading cause of hereditary sensory deaf-blindness in humans; three clinically distinct types of USH have been identified, type 1 to 3. The gene encoding harmonin (USH1C) is the causative gene for the USH type 1C phenotype. There are at least 10 alternatively spliced isoforms of harmonin, which are divided into three subclasses (a, b, and c). All isoforms contain the first two PDZ domains and the first coiled-coil domain. The a and b isoforms all have a third PDZ domain. The different PDZ domains are responsible for interactions with all known Usher syndrome type 1 proteins, and most Usher syndrome type 2 proteins. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This harmonin family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467220 [Multi-domain] Cd Length: 82 Bit Score: 42.69 E-value: 5.64e-05
|
||||||||
| PDZ2_L-delphilin-like | cd06744 | PDZ domain 2 of delphilin (L-delphilin isoform), and related domains; PDZ (PSD-95 ... |
447-488 | 5.87e-05 | ||||
PDZ domain 2 of delphilin (L-delphilin isoform), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of delphilin (also known as glutamate receptor, ionotropic, delta 2-interacting protein 1, L-delphilin). Delphilin, a postsynaptic protein which it is selectively expressed at cerebellar Purkinje cells, links the glutamate receptor delta 2 subunit (GluRdelta2) with the actin cytoskeleton and various signaling molecules. Two alternatively spliced isoforms of delphilin have been characterized: L-delphilin has two PDZ domains, PDZ1 and PDZ2, and S-delphilin has a single PDZ domain (PDZ2). These two isoforms are differently palmitoylated and may be involved in controlling GluRdelta2 signaling in Purkinje cells. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This delphilin-like family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F Pssm-ID: 467226 [Multi-domain] Cd Length: 75 Bit Score: 42.26 E-value: 5.87e-05
|
||||||||
| DegQ | COG0265 | Periplasmic serine protease, S1-C subfamily, contain C-terminal PDZ domain [Posttranslational ... |
446-490 | 6.02e-05 | ||||
Periplasmic serine protease, S1-C subfamily, contain C-terminal PDZ domain [Posttranslational modification, protein turnover, chaperones]; Pssm-ID: 440035 [Multi-domain] Cd Length: 274 Bit Score: 45.91 E-value: 6.02e-05
|
||||||||
| vRING-HC-C4C4_RBBP6 | cd16620 | Variant RING finger, HC subclass (C4C4-type), found in retinoblastoma-binding protein 6 (RBBP6) ... |
13-42 | 6.32e-05 | ||||
Variant RING finger, HC subclass (C4C4-type), found in retinoblastoma-binding protein 6 (RBBP6) and similar proteins; RBBP6, also known as proliferation potential-related protein, protein P2P-R, retinoblastoma-binding Q protein 1 (RBQ-1), or p53-associated cellular protein of testis (PACT), is a nuclear E3 ubiquitin-protein ligase involved in multiple processes, such as the control of gene expression, mitosis, cell differentiation, and cell apoptosis. It plays a role in both promoting and inhibiting apoptosis in many human cancers, including esophageal, lung, hepatocellular, and colon cancers, familial myeloproliferative neoplasms, as well as in human immunodeficiency virus-associated nephropathy (HIVAN). It functions as an Rb- and p53-binding protein that plays an important role in chaperone-mediated ubiquitination and possibly in protein quality control. It acts as a scaffold protein to promote the assembly of the p53/TP53-MDM2 complex, resulting in an increase of MDM2-mediated ubiquitination and degradation of p53/TP53, and leading to both apoptosis and cell growth. It is also a double-stranded RNA-binding protein that plays a role in mRNA processing by regulating the human polyadenylation machinery and modulating expression of mRNAs with AU-rich 3' untranslated regions (UTRs). Moreover, RBBP6 ubiquitinates and destabilizes the transcriptional repressor ZBTB38 that negatively regulates transcription and levels of the MCM10 replication factor on chromatin. Furthermore, RBBP6 is involved in tunicamycin-induced apoptosis by mediating protein kinase (PKR) activation. RBBP6 contains an N-terminal ubiquitin-like domain and a C4C4-type RING finger, whose overall folding is similar to that of the typical C3HC4-type RING-HC finger. RBBP6 interacts with chaperones Hsp70 and Hsp40 through its N-terminal ubiquitin-like domain. It promotes the ubiquitination of p53 by Hdm2 in an E4-like manner through its RING finger. It also interacts directly with the pro-proliferative transcription factor Y-box-binding protein-1 (YB-1) via its RING finger. Pssm-ID: 438282 [Multi-domain] Cd Length: 55 Bit Score: 41.62 E-value: 6.32e-05
|
||||||||
| RING-HC_TRIM62_C-IV | cd16608 | RING finger, HC subclass, found in tripartite motif-containing protein 62 (TRIM62) and similar ... |
15-57 | 7.00e-05 | ||||
RING finger, HC subclass, found in tripartite motif-containing protein 62 (TRIM62) and similar proteins; TRIM62, also known as Ductal Epithelium Associated Ring Chromosome 1 (DEAR1), is a cytoplasmic E3 ubiquitin-protein ligase that was identified as a dominant regulator of acinar morphogenesis in the mammary gland. It is implicated in the inflammatory response of immune cells by regulating the Toll-like receptor 4 (TLR4) signaling pathway, leading to increased activity of the activator protein 1 (AP-1) transcription factor in primary macrophages. It is also involved in muscular protein homeostasis, especially during inflammation-induced atrophy, and may play a role in the pathogenesis of ICU-acquired weakness (ICUAW) by activating and maintaining inflammation in myocytes. Moreover, TRIM62 facilitates K27-linked poly-ubiquitination of CARD9 and also regulates CARD9-mediated anti-fungal immunity and intestinal inflammation. It also functions as a chromosome 1p35 tumor suppressor and negatively regulates transforming growth factor beta (TGFbeta)-driven epithelial-mesenchymal transition (EMT) by binding to and promoting the ubiquitination of SMAD3, a major effector of TGFbeta-mediated EMT. TRIM62 belongs to the C-IV subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region, as well as a B30.2/SPRY (SplA and ryanodine receptor) domain positioned C-terminal to the RBCC domain. Pssm-ID: 438270 [Multi-domain] Cd Length: 52 Bit Score: 41.33 E-value: 7.00e-05
|
||||||||
| PDZ6_GRIP1-2-like | cd06683 | PDZ domain 6 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related ... |
431-493 | 7.56e-05 | ||||
PDZ domain 6 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR) binding proteins GRIP1 (ABP/GRIP2) and GRIP2, and related domains. GRIP1 and GRIP2 each have 7 PDZ domains. The interaction of GRIP1 and GRIP2 with GluA2/3 (AMPAR subunit) regulates AMPAR trafficking and synaptic targeting. GRIP1 has an essential role in regulating AMPAR trafficking during synaptic plasticity and learning and memory. GRIP1 and GRIP2 interact with a variety of other proteins associated with protein trafficking and internalization, for example GRIP1 also interacts with KIF5 (also known as kinesin 1), EphB receptors, scaffold protein liprin-alpha, and the rasGEF GRASP-1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This GRIP family PDZ6 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467171 [Multi-domain] Cd Length: 85 Bit Score: 42.29 E-value: 7.56e-05
|
||||||||
| PDZ5_GRIP1-2-like | cd06682 | PDZ domain 5 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related ... |
431-492 | 7.71e-05 | ||||
PDZ domain 5 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR) binding proteins GRIP1 (ABP/GRIP2) and GRIP2, and related domains. GRIP1 and GRIP2 each have 7 PDZ domains. The interaction of GRIP1 and GRIP2 with GluA2/3 (AMPAR subunit) regulates AMPAR trafficking and synaptic targeting. GRIP1 has an essential role in regulating AMPAR trafficking during synaptic plasticity and learning and memory. GRIP1 and GRIP2 interact with a variety of other proteins associated with protein trafficking and internalization, for example GRIP1 also interacts with KIF5 (also known as kinesin 1), EphB receptors, scaffold protein liprin-alpha, and the rasGEF GRASP-1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This GRIP family domain PDZ5 is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467170 [Multi-domain] Cd Length: 85 Bit Score: 42.33 E-value: 7.71e-05
|
||||||||
| RING-H2_AMFR | cd16455 | RING finger, H2 subclass, found in autocrine motility factor receptor (AMFR) and similar ... |
17-60 | 7.79e-05 | ||||
RING finger, H2 subclass, found in autocrine motility factor receptor (AMFR) and similar proteins; AMFR, also known as AMF receptor, or RING finger protein 45, or ER-protein gp78, is an internalizing cell surface glycoprotein localized in both plasma membrane caveolae and the endoplasmic reticulum (ER). It is involved in the regulation of cellular adhesion, proliferation, motility and apoptosis, as well as in the process of learning and memory. AMFR also functions as a RING finger-dependent ubiquitin protein ligase (E3) implicated in the degradation from the ER. AMFR contains an N-terminal RING-H2 finger and a C-terminal ubiquitin-associated (UBA)-like CUE domain. Pssm-ID: 438119 [Multi-domain] Cd Length: 44 Bit Score: 40.89 E-value: 7.79e-05
|
||||||||
| RING-HC_TRIM43-like_C-IV | cd16603 | RING finger, HC subclass, found in tripartite motif-containing proteins TRIM43, TRIM48, TRIM49, ... |
15-56 | 8.15e-05 | ||||
RING finger, HC subclass, found in tripartite motif-containing proteins TRIM43, TRIM48, TRIM49, TRIM51, TRIM64 and similar proteins; The family includes a group of closely related uncharacterized tripartite motif-containing proteins, TRIM43, TRIM43B, TRIM48/RNF101, TRIM49/RNF18, TRIM49B, TRIM49C/TRIM49L2, TRIM49D/TRIM49L, TRIM51/SPRYD5, TRIM64, TRIM64B, and TRIM64C, whose biological function remain unclear. TRIM49, also known as testis-specific RING-finger protein, has moderate similarity with SS-A/Ro52 antigen, suggesting it may be one of the target proteins of autoantibodies in the sera of patients with these autoimmune disorders. All family members belong to the C-IV subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains as well as a SPRY/B30.2 domain positioned C-terminal to the RBCC domain. In RBCC region, they all have a C3HC4-type RING-HC finger. Pssm-ID: 438265 [Multi-domain] Cd Length: 59 Bit Score: 41.31 E-value: 8.15e-05
|
||||||||
| cpPDZ2_EcRseP-like | cd23083 | circularly permuted PDZ domain 2 (PDZ-C) of Escherichia coli Regulator of sigma-E protease ... |
287-351 | 8.45e-05 | ||||
circularly permuted PDZ domain 2 (PDZ-C) of Escherichia coli Regulator of sigma-E protease (RseP) and related domains; Permuted PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of ResP (also known as Site-2 protease RseP, and YaeL) and related domains. RseP is involved in the regulation of an extracytoplasmic stress response through the cleavage of membrane-spanning anti-stress-response transcription factor (anti-sigmaE) protein RseA; it cleaves the peptide bond between the critical alanine and cysteine in the transmembrane region of RseA, releasing the cytoplasmic domain of RseA with it associated sigmaE. RseP contains two tandem-arranged periplasmic PDZ domains (PDZ-N/PDZ1 and PDZ-C/PDZ2) which act to negatively regulate protease action on intact RseA; they serve as a size-exclusion filter which prevents the access of an intact RseA into the active site of RseP. PDZ domains usually bind in sequence-specific manner to short peptide sequences located at the C-terminal of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains and as well as those with circular permutations and domain swapping of beta-strands. The canonical PDZ domain contains six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. This RseP family PDZ domain is a circularly permuted PDZ domain which places both beta-strands A and B at the C-terminus. Another permutation exists in the PDZ superfamily which places beta-strand A at the C-terminus. Pssm-ID: 467640 [Multi-domain] Cd Length: 85 Bit Score: 42.11 E-value: 8.45e-05
|
||||||||
| DegQ | COG0265 | Periplasmic serine protease, S1-C subfamily, contain C-terminal PDZ domain [Posttranslational ... |
276-337 | 8.46e-05 | ||||
Periplasmic serine protease, S1-C subfamily, contain C-terminal PDZ domain [Posttranslational modification, protein turnover, chaperones]; Pssm-ID: 440035 [Multi-domain] Cd Length: 274 Bit Score: 45.53 E-value: 8.46e-05
|
||||||||
| RING-H2_RNF145 | cd16684 | RING finger, H2 subclass, found in RING finger protein 145 (RNF145) and similar proteins; ... |
18-53 | 1.05e-04 | ||||
RING finger, H2 subclass, found in RING finger protein 145 (RNF145) and similar proteins; RNF145 is an uncharacterized RING finger protein encoded by the RNF145 gene, which is expressed in T lymphocytes, and its expression is altered in acute myelomonocytic and acute promyelocytic leukemias. Although its biological function remains unclear, RNF145 shows high sequence similarity with RNF139, an endoplasmic reticulum (ER)-resident multi-transmembrane protein that functions as a potent growth suppressor in mammalian cells, inducing G2/M arrest, decreased DNA synthesis and increased apoptosis. Like RNF139, RNF145 contains a C3H2C3-type RING-H2 finger with possible E3-ubiquitin ligase activity. Pssm-ID: 319598 [Multi-domain] Cd Length: 43 Bit Score: 40.81 E-value: 1.05e-04
|
||||||||
| RING-HC_RNF113A_B | cd16539 | RING finger, HC subclass, found in RING finger proteins RNF113A, RNF113B, and similar proteins; ... |
13-43 | 1.08e-04 | ||||
RING finger, HC subclass, found in RING finger proteins RNF113A, RNF113B, and similar proteins; RNF113A, also known as zinc finger protein 183 (ZNF183), is an E3 ubiquitin-protein ligase that physically interacts with the E2 protein, UBE2U. A nonsense mutation in RNF113A is associated with an X-linked trichothiodystrophy (TTD). Its yeast ortholog Cwc24p is predicted to have a spliceosome function and acts in a complex with Cef1p to participate in pre-U3 snoRNA splicing, indirectly affecting pre-rRNA processing. It is also important for the U2 snRNP binding to primary transcripts and co-migrates with spliceosomes. Moreover, the ortholog of RNF113A in fruit flies may also act as a spliceosome and is hypothesized to be involved in splicing, namely within the central nervous system. The ortholog in Caenorhabditis elegans is involved in DNA repair of inter-strand crosslinks. RNF113B, also known as zinc finger protein 183-like 1, shows high sequence similarity with RNF113A. Both RNF113A and RNF113B contain a CCCH-type zinc finger, which is commonly found in RNA-binding proteins involved in splicing, and a C3HC4-type RING-HC finger, which is frequently found in E3 ubiquitin ligases. Pssm-ID: 438201 [Multi-domain] Cd Length: 54 Bit Score: 41.04 E-value: 1.08e-04
|
||||||||
| PDZ3_PDZD7-like | cd06751 | PDZ domain 3 of the canonical isoform 1 of PDZ domain containing 7 (PDZD7), and related ... |
248-336 | 1.18e-04 | ||||
PDZ domain 3 of the canonical isoform 1 of PDZ domain containing 7 (PDZD7), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 3 of the long isoform 1 of PDZD7, and related domains. PDZD7 is critical for the organization of the Usher syndrome type 2 (USH2) complex. Usher syndrome is the leading cause of hereditary sensory deaf-blindness in humans; USH2 is the most common sub-type. Formation of the USH2 complex is based upon heterodimerization between PDZD7 and whirlin (another PDZ domain-containing protein) and a subsequent dynamic interplay between USH2 proteins via their multiple PDZ domains. The PDZD7 PDZ2 domain binds GPR98 (also known as VLGR1) and usherin (USH2A). PDZD7 and whirlin form heterodimers through their multiple PDZ domains; whirlin and PDZD7 interact with usherin and GPR98 to form an interdependent ankle link complex. PDZD7 also interacts with myosin VIIa and can also form homodimers through its PDZ2 domain. Various isoforms of PDZD7 produced by alternative splicing have been identified; this subgroup includes the third PDZ domain of the canonical isoform of PDZD7- isoform 1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDZD7-like family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467233 [Multi-domain] Cd Length: 89 Bit Score: 42.04 E-value: 1.18e-04
|
||||||||
| RAD18 | COG5432 | RING-finger-containing E3 ubiquitin ligase [Signal transduction mechanisms]; |
12-57 | 1.28e-04 | ||||
RING-finger-containing E3 ubiquitin ligase [Signal transduction mechanisms]; Pssm-ID: 227719 [Multi-domain] Cd Length: 391 Bit Score: 45.46 E-value: 1.28e-04
|
||||||||
| RING-HC_TRIM65_C-IV | cd16609 | RING finger, HC subclass, found in tripartite motif-containing protein TRIM65 and similar ... |
15-57 | 1.35e-04 | ||||
RING finger, HC subclass, found in tripartite motif-containing protein TRIM65 and similar proteins; TRIM65 is an E3 ubiquitin-protein ligase that interacts with the innate immune receptor MDA5, enhancing its ability to stimulate interferon-beta signaling. It functions as a potential oncogenic protein that negatively regulates p53 through ubiquitination, providing insight into the development of novel approaches targeting TRIM65 for non-small cell lung carcinoma (NSCLC) treatment, and also overcoming chemotherapy resistance. Moreover, TRIM65 negatively regulates microRNA-driven suppression of mRNA translation by targeting TNRC6 proteins for ubiquitination and degradation. TRIM65 belongs to the C-IV subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region, as well as a B30.2/SPRY (SplA and ryanodine receptor) domain positioned C-terminal to the RBCC domain. Pssm-ID: 438271 [Multi-domain] Cd Length: 58 Bit Score: 40.82 E-value: 1.35e-04
|
||||||||
| cpPDZ_EcRseP-like | cd23081 | circularly permuted PDZ domains of Escherichia coli Regulator of sigma-E protease (RseP) and ... |
449-502 | 1.37e-04 | ||||
circularly permuted PDZ domains of Escherichia coli Regulator of sigma-E protease (RseP) and related domains; Permuted PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of ResP (also known as Site-2 protease RseP, and YaeL), and related domains. RseP is involved in the regulation of an extracytoplasmic stress response through the cleavage of membrane-spanning anti-stress-response transcription factor (anti-sigmE) protein RseA; it cleaves the peptide bond between the critical alanine and cysteine in the transmembrane region of RseA, releasing the cytoplasmic domain of RseA with its associated sigmaE. RseP contains two tandem-arranged periplasmic PDZ domains (PDZ-N/PDZ1 and PDZ-C/PDZ2) which act to negatively regulate protease action on intact RseA; they serve as a size-exclusion filter which prevents the access of an intact RseA into the active site of RseP. PDZ domains usually bind in sequence-specific manner to short peptide sequences located at the C-terminal of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains and as well as those with circular permutations and domain swapping of beta-strands. The canonical PDZ domain contains six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. This RseP family PDZ domain is a circularly permuted PDZ domain which places both beta-strands A and B at the C-terminus. Another permutation exists in the PDZ superfamily which places beta-strand A at the C-terminus. Pssm-ID: 467638 [Multi-domain] Cd Length: 83 Bit Score: 41.41 E-value: 1.37e-04
|
||||||||
| RING-HC_Topors | cd16574 | RING finger, HC subclass, found in topoisomerase I-binding arginine/serine-rich protein ... |
15-59 | 1.38e-04 | ||||
RING finger, HC subclass, found in topoisomerase I-binding arginine/serine-rich protein (Topors) and similar proteins; Topors, also known as topoisomerase I-binding RING finger protein, tumor suppressor p53- binding protein 3, or p53-binding protein 3 (p53BP3), is a ubiquitously expressed nuclear E3 ubiquitin-protein ligase that can ligate both ubiquitin and small ubiquitin-like modifier (SUMO) to substrate proteins in the nucleus. It contains an N-terminal C3HC4-type RING-HC finger which ligates ubiquitin to its target proteins including DNA topoisomerase I, p53, NKX3.1, H2AX, and the AAV-2 Rep78/68 proteins. As a RING-dependent E3 ubiquitin ligase, Topors works with the E2 enzymes UbcH5a, UbcH5c, and UbcH6, but not with UbcH7, CDC34, or UbcH2b. Topors acts as a tumor suppressor in various malignancies. It regulates p53 modification, suggesting it may be responsible for astrocyte elevated gene-1 (AEG-1, also known as metadherin, or LYRIC) ubiquitin modification. Plk1-mediated phosphorylation of Topors inhibits Topors-mediated sumoylation of p53, whereas p53 ubiquitination is enhanced, leading to p53 degradation. It also functions as a negative regulator of the prostate tumor suppressor NKX3.1. Moreover, Topors is associated with promyelocytic leukemia nuclear bodies, and may be involved in the cellular response to camptothecin. It also plays a key role in the turnover of H2AX protein, discriminating the type of DNA damaging stress. Furthermore, Topors is a cilia-centrosomal protein associated with autosomal dominant retinal degeneration. Mutations in TOPORS cause autosomal dominant retinitis pigmentosa (adRP). Pssm-ID: 438236 [Multi-domain] Cd Length: 47 Bit Score: 40.35 E-value: 1.38e-04
|
||||||||
| PDZ_PDLIM-like | cd06753 | PDZ domain of PDZ-LIM family proteins, and related domains; PDZ (PSD-95 (Postsynaptic density ... |
260-337 | 1.53e-04 | ||||
PDZ domain of PDZ-LIM family proteins, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of PDZ-LIM family proteins including PDLIM1-7, and related domains. PDZ-LIM family proteins (also known as Zasp PDZ domain proteins) are involved in the rearrangement of the actin cytoskeleton; they mediate association with the cytoskeleton through alpha-actinin as well as with other proteins involved in signal transduction pathways. Members of this family include PDLIM1 (also known as C-terminal LIM domain protein 1, elfin, LIM domain protein CLP-36), PDLIM2 (also known as PDZ-LIM protein mystique), PDLIM3 (also known as actinin-associated LIM protein, alpha-actinin-2-associated LIM protein, ALP), PDLIM4 (also known as LIM protein RIL, Reversion-induced LIM protein), PDLIM5 (also known as enigma homolog, ENH, enigma-like PDZ and LIM domains protein), PDLIM6 (also known as LIM domain-binding protein 3, ZASP, Cypher, Oracle), and PDLIM7 (also known as PDZ and LIM domain protein 7, LIM mineralization protein, LMP; protein enigma). PDLIM1 has been shown to negatively regulate NF-kappaB-mediated signaling in the cytoplasm. PDLIM7 negatively regulates p53 through binding murine double minute 2 (MDM2). The PDZ domains of PDZ-LIM family proteins PDLIM1, 2, 3, 5, 6, 7 have been shown to bind actin. Other PDZ-LIM family PDZ domain binding partners include thyroid receptor interacting protein-6 (PDLIM4-PDZ), the LIM domain of PDLIM4 (PDLIM4-PDZ), tropomyosin (PDLIM7-PDZ), myotilin and calsarcin 1 (PDLIM6-PDZ), and proteins from the myotilin and FATZ (calsarcin/myozenin) families (PDLIM1, 3, 4, 6 PDZ domains). PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PDLIM-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467235 [Multi-domain] Cd Length: 79 Bit Score: 41.36 E-value: 1.53e-04
|
||||||||
| PDZ_ARHGEF11-12-like | cd23069 | PDZ domain of ARHGEF11, ARHGEF12, and related domains; PDZ (PSD-95 (Postsynaptic density ... |
428-499 | 1.63e-04 | ||||
PDZ domain of ARHGEF11, ARHGEF12, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of ARHGEF11, ARHGEF12, and related domains. This subfamily includes the GEFs (guanine exchange factors) ARHGEF11 (Rho guanine nucleotide exchange factor 11, known as PDZ-RhoGEF) and ARHGEF12 (Rho guanine nucleotide exchange factor 12, also known as leukemia-associated RhoGEF). GEFs activate Rho GTPases by promoting GTP binding. ARHGEF11/12 are regulators of G protein signaling (RGS) domain-containing GEFs; the RGS domain mediates their binding to and activation of Galpha (and Gq also in the case of ARHGEF12), in response to G-protein coupled receptor activation. ARHGEF11 and 12 are involved in serum-signaling, and regulate Yes-Associated Protein (YAP1)-dependent transcription. The ARHGEF12 PDZ domain binds plexin-B1 and the receptor tyrosine kinase insulin-like growth factor receptor (IGF-R1) beta-subunit. ARHGEF12 also interacts with glutamate receptor delta-1(GluD1), a postsynaptic organizer of inhibitory synapses in cortical pyramidal neurons. The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This ARHGEF11-12-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467282 [Multi-domain] Cd Length: 76 Bit Score: 41.22 E-value: 1.63e-04
|
||||||||
| mRING-HC-C4C4_TRIM37_C-VIII | cd16619 | Modified RING finger, HC subclass (C4C4-type), found in tripartite motif-containing protein 37 ... |
16-57 | 1.64e-04 | ||||
Modified RING finger, HC subclass (C4C4-type), found in tripartite motif-containing protein 37 (TRIM37) and similar proteins; TRIM37, also known as mulibrey nanism protein, or MUL, is a peroxisomal E3 ubiquitin-protein ligase that is involved in the tumorigenesis of several cancer types, including pancreatic ductal adenocarcinoma (PDAC), hepatocellular carcinoma (HCC), breast cancer, and sporadic fibrothecoma. It mono-ubiquitinates histone H2A, a chromatin modification associated with transcriptional repression. Moreover, TRIM37 possesses anti-HIV-1 activity, and interferes with viral DNA synthesis. Mutations in the human TRIM37 gene (also known as MUL) cause Mulibrey (muscle-liver-brain-eye) nanism, a rare growth disorder of prenatal onset characterized by dysmorphic features, pericardial constriction, and hepatomegaly. TRIM37 belongs to the C-VIII subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C4C4-type RING finger, whose overall folding is similar to that of the typical C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region, as well as a MATH (meprin and TRAF-C homology) domain positioned C-terminal to the RBCC domain. Its MATH domain has been shown to interact with the TRAF (TNF-Receptor-Associated Factor) domain of six known TRAFs in vitro. Pssm-ID: 438281 [Multi-domain] Cd Length: 43 Bit Score: 40.03 E-value: 1.64e-04
|
||||||||
| PDZ_PICK1-like | cd06722 | PDZ domain of PICK1 (protein interacting with C-kinase 1) and similar domains; PDZ (PSD-95 ... |
249-334 | 1.90e-04 | ||||
PDZ domain of PICK1 (protein interacting with C-kinase 1) and similar domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of PICK1, and related domains. PICK1 (also known as PRKCA-binding protein and protein kinase C-alpha-binding protein) plays a key role in regulating trafficking of binding partners by altering either their subcellular targeting and/or surface expression. PICK1 plays a role in synaptic plasticity by regulating the trafficking and internalization of amino-3-hydroxy-5-methylisoxazole-4-propionic acid (AMPA) receptors; the PICK1-PDZ domain binds the AMPA receptor subunits. The PICK1 PDZ domain also binds glutamate transporters, Eph receptors, metabotropic glutamate receptors, and ASICs (acid-sensing ion channels), among others. Clustering and synaptic targeting of PICK1 requires direct interaction between the PDZ domain and lipid membranes. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PICK-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged as beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta- strand F. Pssm-ID: 467205 [Multi-domain] Cd Length: 84 Bit Score: 41.25 E-value: 1.90e-04
|
||||||||
| zf-C3HC4_4 | pfam15227 | zinc finger of C3HC4-type, RING; This is a family of primate-specific Ret finger protein-like ... |
18-45 | 1.91e-04 | ||||
zinc finger of C3HC4-type, RING; This is a family of primate-specific Ret finger protein-like (RFPL) zinc-fingers of the C3HC4 type. Ret finger protein-like proteins are primate-specific target genes of Pax6, a key transcription factor for pancreas, eye and neocortex development. This domain is likely to be DNA-binding. This zinc-finger domain together with the RDM domain, pfam11002, forms a large zinc-finger structure of the RING/U-Box superfamily. RING-containing proteins are known to exert an E3 ubiquitin protein ligase activity with the zinc-finger structure being mandatory for binding to the E2 ubiquitin-conjugating enzyme. Pssm-ID: 464570 [Multi-domain] Cd Length: 42 Bit Score: 39.73 E-value: 1.91e-04
|
||||||||
| RING-H2 | cd16448 | H2 subclass of RING (RING-H2) fingers and its variants; The RING finger is a specialized type ... |
18-57 | 1.92e-04 | ||||
H2 subclass of RING (RING-H2) fingers and its variants; The RING finger is a specialized type of Zn-finger of 40 to 60 residues that binds two atoms of zinc. It is defined by the "cross-brace" motif that chelates zinc atoms by eight amino acid residues, typically Cys or His, arranged in a characteristic spacing. Canonical RING motifs have been categorized into two major subclasses, RING-HC (C3HC4-type) and RING-H2 (C3H2C3-type), according to their Cys/His content. There are also many variants of RING fingers: some have different Cys/His patterns while some lack a single Cys or His residue at typical Zn ligand positions (the fourth or eighth zinc ligand is prevalently exchanged for an Asp, which can indeed chelate Zn in a RING finger as well). This family corresponds to the H2 subclass of RING (RING-H2) finger proteins that are characterized by containing C3H2C3-type canonical RING-H2 fingers or noncanonical RING-H2 finger variants, including C4HC3- (RING-CH alias RINGv), C3H3C2-, C3H2C2D-, C3DHC3-, and C4HC2H-type modified RING-H2 fingers. The canonical RING-H2 finger has been defined as C-X2-C-X(9-39)-C-X(1-3)-H-X(2-3)-H-X2-C-X(4-48)-C-X2-C, X is any amino acid and the number of X residues varies in different fingers. It binds two Zn ions in a unique "cross-brace" arrangement, which distinguishes it from tandem zinc fingers and other similar motifs. RING-H2 finger can be found in a group of diverse proteins with a variety of cellular functions, including oncogenesis, development, viral replication, signal transduction, the cell cycle and apoptosis. Many of them are ubiquitin-protein ligases (E3s) that serves as a scaffold for binding to ubiquitin-conjugating enzymes (E2s, also referred to as ubiquitin carrier proteins or UBCs) in close proximity to substrate proteins, which enables efficient transfer of ubiquitin from E2 to the substrates. Pssm-ID: 438112 [Multi-domain] Cd Length: 43 Bit Score: 39.69 E-value: 1.92e-04
|
||||||||
| PDZ1_harmonin | cd06737 | PDZ domain 1 of harmonin isoforms a, b, and c, and related domains; PDZ (PSD-95 (Postsynaptic ... |
426-490 | 1.94e-04 | ||||
PDZ domain 1 of harmonin isoforms a, b, and c, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of harmonin isoforms a, b, and c, and related domains. Harmonin (also known as Usher Type 1C, PDZ-73 and AIE-75) is a key organizer of the Usher (USH) protein interactome. USH syndrome is the leading cause of hereditary sensory deaf-blindness in humans; three clinically distinct types of USH have been identified, type 1 to 3. The gene encoding harmonin (USH1C) is the causative gene for the USH type 1C phenotype. There are at least 10 alternatively spliced isoforms of harmonin, which are divided into three subclasses (a, b, and c). All isoforms contain the first two PDZ domains and the first coiled-coil domain. The a and b isoforms all have a third PDZ domain. The different PDZ domains are responsible for interactions with all known Usher syndrome type 1 proteins, and most Usher syndrome type 2 proteins. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This harmonin family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467219 [Multi-domain] Cd Length: 85 Bit Score: 41.09 E-value: 1.94e-04
|
||||||||
| RING-HC_TRIM4_C-IV | cd16590 | RING finger, HC subclass, found in tripartite motif-containing protein TRIM4 and similar ... |
11-57 | 1.96e-04 | ||||
RING finger, HC subclass, found in tripartite motif-containing protein TRIM4 and similar proteins; TRIM4 belongs to the C-IV subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox2, and a coiled coil region, as well as a SPRY/B30.2 domain positioned C-terminal to the RBCC domain. TRIM4, also known as RING finger protein 87 (RNF87), is a cytoplasmic E3 ubiquitin-protein ligase that has recently evolved and is present only in higher mammals. It transiently interacts with mitochondria, induces mitochondrial aggregation and sensitizes the cells to hydrogen peroxide (H2O2) induced death. Its interaction with peroxiredoxin 1 (PRX1) is critical for the regulation of H2O2 induced cell death. Moreover, TRIM4 functions as a positive regulator of RIG-I-mediated type I interferon induction. It regulates the K63-linked ubiquitination of RIG-1 and assembly of antiviral signaling complex at the mitochondria. Pssm-ID: 438252 [Multi-domain] Cd Length: 61 Bit Score: 40.40 E-value: 1.96e-04
|
||||||||
| RING-HC_RAG1 | cd16530 | RING finger, HC subclass, found in recombination activating gene-1 (RAG-1) and similar ... |
18-57 | 2.09e-04 | ||||
RING finger, HC subclass, found in recombination activating gene-1 (RAG-1) and similar proteins; RAG-1, also known as V(D)J recombination-activating protein 1, RING finger protein 74 (RNF74), or endonuclease RAG1, is the catalytic component of the RAG complex, a multiprotein complex that mediates the DNA cleavage phase during V(D)J recombination. RAG1 is a lymphoid-specific factor that mediates DNA-binding to conserved recombination signal sequences (RSS) and catalyzes DNA cleavage activities by introducing a double-strand break between the RSS and the adjacent coding segment. It also functions as an E3 ubiquitin-protein ligase that mediates monoubiquitination of histone H3, which is required for the joining step of V(D)J recombination. RAG-1 contains an N-terminal C3HC4-type RING-HC finger that mediates monoubiquitylation of histone H3, an adjacent C2H2-type zinc finger, and a nonamer binding (NBD) DNA-binding domain. Pssm-ID: 319444 [Multi-domain] Cd Length: 46 Bit Score: 39.73 E-value: 2.09e-04
|
||||||||
| PDZ_2 | pfam13180 | PDZ domain; |
280-338 | 2.16e-04 | ||||
PDZ domain; Pssm-ID: 433015 [Multi-domain] Cd Length: 74 Bit Score: 40.72 E-value: 2.16e-04
|
||||||||
| zf-C3HC4 | pfam00097 | Zinc finger, C3HC4 type (RING finger); The C3HC4 type zinc-finger (RING finger) is a ... |
18-53 | 2.21e-04 | ||||
Zinc finger, C3HC4 type (RING finger); The C3HC4 type zinc-finger (RING finger) is a cysteine-rich domain of 40 to 60 residues that coordinates two zinc ions, and has the consensus sequence: C-X2-C-X(9-39)-C-X(1-3)-H-X(2-3)-C-X2-C-X(4-48)-C-X2-C where X is any amino acid. Many proteins containing a RING finger play a key role in the ubiquitination pathway. Pssm-ID: 395049 [Multi-domain] Cd Length: 40 Bit Score: 39.65 E-value: 2.21e-04
|
||||||||
| RING-HC_AtBRCA1-like | cd23147 | RING finger, HC subclass, found in Arabidopsis thaliana protein BREAST CANCER SUSCEPTIBILITY 1 ... |
15-53 | 2.24e-04 | ||||
RING finger, HC subclass, found in Arabidopsis thaliana protein BREAST CANCER SUSCEPTIBILITY 1 homolog (AtBRCA1) and similar proteins; AtBRCA1 plays a role in DNA repair and in cell-cycle control. It is required for the repair of DNA double-strand breaks (DSBs), both natural and induced by genotoxic stress, by homologous recombination (HR). AtBRCA1 contains a typical C3HC4-type RING-HC finger. Pssm-ID: 438509 [Multi-domain] Cd Length: 54 Bit Score: 40.14 E-value: 2.24e-04
|
||||||||
| mRING-HC-C3HC3D_TRAF5 | cd16642 | Modified RING finger, HC subclass (C3HC3D-type), found in tumor necrosis factor (TNF) ... |
17-43 | 2.44e-04 | ||||
Modified RING finger, HC subclass (C3HC3D-type), found in tumor necrosis factor (TNF) receptor-associated factor 5 (TRAF5) and similar proteins; TRAF5, also known as RING finger protein 84 (RNF84), is an important signal transducer for a wide range of TNF receptor superfamily members, including tumor necrosis factor receptor 1 (TNFR1), TNFR2, CD40, and other lymphocyte costimulatory receptors, RANK/TRANCE-R, ectodysplasin-A Receptor (EDAR), lymphotoxin-beta receptor (LT-betaR), latent membrane protein 1 (LMP1), and IRE1. It functions as an activator of NF-kappaB, MAPK, and JNK, and is involved in both RANKL- and TNFalpha-induced osteoclastogenesis. It mediates CD40 signaling by associating with the cytoplasmic tail of CD40. It also negatively regulates Toll-like receptor (TLR) signaling and functions as a negative regulator of the interleukin 6 (IL-6) receptor signaling pathway that limits the differentiation of inflammatory CD4(+) T cells. TRAF5 contains an N-terminal domain with a modified C3HC3D-type RING-HC finger and several zinc fingers, and a C-terminal TRAF domain that comprises a coiled coil domain and a conserved TRAF-C domain. Pssm-ID: 438304 [Multi-domain] Cd Length: 56 Bit Score: 40.11 E-value: 2.44e-04
|
||||||||
| PDZ1_hSTXBP4-PDZ2_GgSTXBP4-like | cd06698 | PDZ1 domain of human syntaxin-binding protein 4 (STXBP4), PDZ2 domain of Gallus gallus ... |
431-489 | 2.47e-04 | ||||
PDZ1 domain of human syntaxin-binding protein 4 (STXBP4), PDZ2 domain of Gallus gallus uncharacterized STXBP4 isoform X1, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of human syntaxin-binding protein 4 (STXBP4), PDZ2 domain of Gallus gallus uncharacterized STXBP4 isoform X1, and related domains. Human STXBP4 (also known as Synip) includes a single PDZ domain, a coiled-coil domain, and a WW domain (named for its two conserved tryptophans); Gallus gallus STXBP4 isoform X1 contains 2 PDZ domains (PDZ1 and PDZ2). Human STXBP4 plays a role in the translocation of transport vesicles from the cytoplasm to the plasma membrane: insulin induces the dissociation of the STXBP4 and STX4 complex liberating STX4 to interact with Vamp2, and to form the SNARE complex thereby promoting vesicle fusion. It may also play a role in the regulation of insulin release by pancreatic beta cells after stimulation by glucose. Human STXBP4 is also known to physically associate with a prominent isoform of TP63 (deltaNp63alpha 9) whose overexpression promotes squamous cell carcinoma development, and in doing so prevents degradation of this isoform by the Cdc20-APC/C complex, Itch, and RACK1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This STXBP4-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467184 [Multi-domain] Cd Length: 89 Bit Score: 41.14 E-value: 2.47e-04
|
||||||||
| Peptidase_M50 | pfam02163 | Peptidase family M50; |
448-502 | 2.49e-04 | ||||
Peptidase family M50; Pssm-ID: 426630 [Multi-domain] Cd Length: 291 Bit Score: 44.41 E-value: 2.49e-04
|
||||||||
| PDZ_FRMPD1_3_4-like | cd06769 | PDZ domain of FERM and PDZ domain-containing protein 1 (FRMPD1), FRMPD3, FRMPD4, and related ... |
249-336 | 2.79e-04 | ||||
PDZ domain of FERM and PDZ domain-containing protein 1 (FRMPD1), FRMPD3, FRMPD4, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of FRMPD1, FRMPD3, FRMPD4, and related domains. FRMPD1 (also known as FERM domain-containing protein 2, FRMD2), inhibits the malignant phenotype of lung cancer by activating the Hippo pathway via interaction with WWC3; the FRMPD1 PDZ domain binds WWC3. FRMPD3 is a target gene of the neuron-specific transcription factor NPAS4 that is involved in synaptic plasticity. FRMPD4 (also known as PDZ domain-containing protein 10, PDZD10, PDZK10, PSD-95-interacting regulator of spine morphogenesis, and Preso) regulates dendritic spine morphogenesis, and mGluR1/5 signaling; the FRMPD4 PDZ domain binds PAK-interacting exchange factor-beta (betaPix). PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This FRMPD1,3,4-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467250 [Multi-domain] Cd Length: 75 Bit Score: 40.31 E-value: 2.79e-04
|
||||||||
| PDZ1_DLG5-like | cd06764 | PDZ domain 1 of Discs Large 5 (Dlg5) and related domains; PDZ (PSD-95 (Postsynaptic density ... |
443-487 | 2.85e-04 | ||||
PDZ domain 1 of Discs Large 5 (Dlg5) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of Drosophila and mammalian Dlg5, and related domains. Dlg5 is a scaffold protein with multiple conserved functions that are independent of each other in regulating growth, cell polarity, and cell adhesion. It has a coiled-coil domain, 4 PDZ domains and a MAGUK domain (an SH3 domain next to a non-catalytically active guanylate kinase domain). Deregulation of Dlg5 has been implicated in the malignancy of several cancer types. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Dlg5-like family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467245 [Multi-domain] Cd Length: 95 Bit Score: 40.85 E-value: 2.85e-04
|
||||||||
| PDZ_SHANK1_3-like | cd06746 | PDZ domain of SH3 and multiple ankyrin repeat domains protein 1 (SHANK1), SHANK2, SHANK3, and ... |
448-488 | 2.96e-04 | ||||
PDZ domain of SH3 and multiple ankyrin repeat domains protein 1 (SHANK1), SHANK2, SHANK3, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of SHANK1, SHANK2, SHANK3, and related domains. SHANK family proteins, SHANK1 (also known as somatostatin receptor-interacting protein, SSTR-interacting protein, SSTRIP), SHANK2 (also known as cortactin-binding protein 1, proline-rich synapse-associated protein 1), and SHANK3 (proline-rich synapse-associated protein 2) are synaptic scaffolding proteins which are highly enriched in the post-synaptic densities of excitatory synapses. They have been implicated in synaptic transmission, synapse formation, synaptic plasticity, and cytoskeletal remodeling, and are regulators of Cav1 calcium current and CREB target expression. Many protein ligands have been identified for the Shank PDZ domain, such as GKAP (also known as SAPAP), betaPIX (a guanine nucleotide exchange factor used by Rho GTPase family members Rac1 and Cdc42), alpha-latrotoxin, neuroligin, group I metabotropic glutamate receptors (mGluRs), and L-type calcium channels. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This SHANK-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged as beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta- strand F. Pssm-ID: 467228 [Multi-domain] Cd Length: 101 Bit Score: 41.04 E-value: 2.96e-04
|
||||||||
| cpPDZ_Deg_HtrA-like | cd06779 | permuted PDZ domain of Deg/high-temperature requirement factor A (HtrA) family of housekeeping ... |
442-488 | 3.11e-04 | ||||
permuted PDZ domain of Deg/high-temperature requirement factor A (HtrA) family of housekeeping serine proteases and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of Deg/HtrA-type serine proteases that participate in folding and degradation of aberrant proteins, and in processing and maturation of native proteins. Typically, these proteases have an N-terminal serine protease domain and at least one C-terminal PDZ domain that recognizes substrates, and in some cases activates the protease function. An exception is yeast Nma11p which has two protease domains and four PDZ domains; its N-terminal half is comprised of a protease domain, followed by two PDZ domains, and its C-terminal half has a similar domain arrangement. HtrA-type proteases include the human HtrA1-4 and MBTPS2, tricorn protease, DegS, DegP and C-terminal processing peptidase, cyanobacterial serine proteases Hhoa, HhoB, and HtrA, and yeast Nma11p. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-termini of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains and as well as those with circular permutations and domain swapping of beta-strands. The canonical PDZ domain contains six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. This Deg/HtrA family PDZ domain is a circularly permuted PDZ domain which places beta-strand A at the C-terminus. Another permutation exists in the PDZ superfamily which places both beta-strands A and B on the C-terminus. Pssm-ID: 467621 [Multi-domain] Cd Length: 91 Bit Score: 40.74 E-value: 3.11e-04
|
||||||||
| PDZ_syntrophin-like | cd06801 | PDZ domain of syntrophins, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), ... |
428-488 | 3.13e-04 | ||||
PDZ domain of syntrophins, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of syntrophins (including alpha-1-syntrophin, beta-1-syntrophin, beta-2-syntrophin, gamma-1-syntrophin, and gamma-2-syntrophin), and related domains. Syntrophins play a role in recruiting various signaling molecules into signaling complexes and help provide appropriate spatiotemporal regulation of signaling pathways. They function in cytoskeletal organization and maintenance; as components of the dystrophin-glycoprotein complex (DGC), they help maintain structural integrity of skeletal muscle fibers. They link voltage-gated sodium channels to the actin cytoskeleton and the extracellular matrix, and control the localization and activity of the actin reorganizing proteins such as PI3K, PI(3,4)P2 and TAPP1. Through association with various cytoskeletal proteins within the cells, they are involved in processes such as regulation of focal adhesions, myogenesis, calcium homeostasis, and cell migration. They also have roles in synapse formation and in the organization of utrophin, acetylcholine receptor, and acetylcholinesterase at the neuromuscular synapse. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This syntrophin-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467262 [Multi-domain] Cd Length: 83 Bit Score: 40.63 E-value: 3.13e-04
|
||||||||
| PDZ_MPP6-MPP2-like | cd10832 | PDZ domain of membrane palmitoylated protein 6 (MPP6), MPP2, and related domains; PDZ (PSD-95 ... |
424-483 | 3.31e-04 | ||||
PDZ domain of membrane palmitoylated protein 6 (MPP6), MPP2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of MPP6, MPP2, and related domains. MPP6 (also known as MAGUK p55 subfamily member, Protein associated with Lin-7, 2 (PALS2), Veli-associated MAGUK 1, and VAM-1) is a membrane-associated guanylate kinase (MAGUK)-like protein. MPP6 is a regulator of Lin-7 expression and localization. MPP6 is also known to bind cell-adhesion protein, nectin-like molecule-2 (Necl-2), and localize to the basolateral plasma membrane in mammalian epithelial cells. MPP2 (also known as MAGUK p55 subfamily member 2) is a postsynaptic protein that links SynCAM1 cell adhesion molecules to core components of the postsynaptic density. Other members of this family include the Drosophila Vari protein, an essential basolateral septate junction protein which interacts with the cell-adhesion protein neurexin IV. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MPP6-MPP2-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467268 [Multi-domain] Cd Length: 78 Bit Score: 40.29 E-value: 3.31e-04
|
||||||||
| PDZ2_MUPP1-like | cd06667 | PDZ domain 2 of multi-PDZ-domain protein 1 (MUPP1) and PATJ (protein-associated tight junction) ... |
445-502 | 3.41e-04 | ||||
PDZ domain 2 of multi-PDZ-domain protein 1 (MUPP1) and PATJ (protein-associated tight junction) and similar domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of MUPP1 and PATJ, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ2 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F Pssm-ID: 467155 [Multi-domain] Cd Length: 80 Bit Score: 40.34 E-value: 3.41e-04
|
||||||||
| PDZ_RIM-like | cd06714 | PDZ domain of Rab3-interacting molecule 1 (RIM), RIM2, piccolo and related domains; PDZ ... |
445-483 | 3.42e-04 | ||||
PDZ domain of Rab3-interacting molecule 1 (RIM), RIM2, piccolo and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of RIM, RIM2, piccolo and related domains. RIM proteins and Gallus gallus protein piccolo (also called aczonin) are involved in neurotransmitter release at presynaptic active zones, the site of vesicle fusion. A protein complex containing RIM proteins positions synaptic vesicles containing synaptotagmin at the active zone. RIM proteins simultaneously activate docking and priming of synaptic vesicles and recruit Ca2+-channels to active zones, thereby connecting primed synaptic vesicles to Ca2+-channels. RIM binding to vesicular Rab proteins (Rab3 and Rab27 isoforms) mediates vesicle docking; RIM binding to Munc13 activates vesicle priming; RIM binding to the Ca2+-channel, both directly and indirectly via RIM-BP, recruits the Ca2+-channels. The RIM PDZ domain interacts with the C-termini of N- and P/Q-type voltage-gated Ca2+-channels. RIM1, RIM2 and piccolo also participate in regulated exocytosis through binding cAMP-GEFII (cAMP-binding protein-guanidine nucleotide exchange factor II). The piccolo PDZ domain binds cAMP-GEFII. RIM2 also plays a role in dendrite formation by melanocytes. Caenorhabditis elegans RIM (also known as unc-10) may be involved in the regulation of defecation and daumone response. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This RIM-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467198 [Multi-domain] Cd Length: 95 Bit Score: 40.61 E-value: 3.42e-04
|
||||||||
| PDZ5_INAD-like | cd23066 | PDZ domain 5 of inactivation no after potential D (INAD), and related domains; PDZ (PSD-95 ... |
280-337 | 3.49e-04 | ||||
PDZ domain 5 of inactivation no after potential D (INAD), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 5 of INAD, and related domains. INAD assembles key enzymes of the Drosophila compound eye photo-transduction pathway into a supramolecular complex, supporting efficient and fast light signaling. It contains 5 PDZ domains arranged in tandem (PDZ1-PDZ5) which independently bind various proteins. INAD PDZ2 binds eye-specific protein kinase C, INAD PDZ3 binds transient receptor potential (TRP) channel, and INAD PDZ45 tandem binds NORPA (phospholipase Cbeta, PLCbeta). Mutations of the inaD gene that lead to disruption of each of these interactions impair fly photo signal transduction. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This INAD-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467279 [Multi-domain] Cd Length: 80 Bit Score: 40.18 E-value: 3.49e-04
|
||||||||
| RING-HC_TRIM60-like_C-IV | cd16607 | RING finger, HC subclass, found in tripartite motif-containing proteins TRIM60, TRIM61, TRIM75 ... |
17-57 | 3.71e-04 | ||||
RING finger, HC subclass, found in tripartite motif-containing proteins TRIM60, TRIM61, TRIM75 and similar proteins; TRIM60, also known as RING finger protein 129 (RNF129) or RING finger protein 33 (RNF33), is a cytoplasmic protein expressed in the testis. It may play an important role in the spermatogenesis process, the development of the preimplantation embryo, and in testicular functions. RNF33 interacts with the cytoplasmic kinesin motor proteins KIF3A and KIF3B suggesting possible contribution to cargo movement along the microtubule in the expressed sites. It is also involved in spermatogenesis in Sertoli cells under the regulation of nuclear factor-kappaB (NF-kappaB). TRIM75 mainly localizes within spindles, suggesting it may function in spindle organization and thereby affect meiosis. Both TRIM60 and TRIM75 belong the C-IV subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, a B2-box, and two coiled coil domains, as well as a PRY domain and a B30.2/SPRY (SplA and ryanodine receptor) domain positioned C-terminal to the RBCC domain. In contrast, TRIM61 belongs to the C-V subclass of the TRIM family that contains RBCC domains only. Its biological function remains unclear. Pssm-ID: 438269 [Multi-domain] Cd Length: 48 Bit Score: 39.33 E-value: 3.71e-04
|
||||||||
| RING-HC_MmTRIM43-like | cd23133 | RING finger, HC subclass, found in Mus musculus tripartite motif-containing protein 43 (TRIM43) ... |
15-56 | 3.71e-04 | ||||
RING finger, HC subclass, found in Mus musculus tripartite motif-containing protein 43 (TRIM43) and similar propteins; This subfamily includes TRIM43A, TRIM43B and TRIM43C, which are expressed specifically in mouse preimplantation embryos. They contain a typical C3HC4-type RING-HC finger. Pssm-ID: 438495 [Multi-domain] Cd Length: 57 Bit Score: 39.51 E-value: 3.71e-04
|
||||||||
| RING-H2_RNF115 | cd16800 | RING finger, H2 subclass, found in RING finger protein 115 (RNF115) and similar proteins; ... |
16-63 | 3.81e-04 | ||||
RING finger, H2 subclass, found in RING finger protein 115 (RNF115) and similar proteins; RNF115, also known as Rab7-interacting ring finger protein (Rabring 7), or zinc finger protein 364 (ZNF364), or breast cancer-associated gene 2 (BCA2), is an E3 ubiquitin-protein ligase that is an endogenous inhibitor of adenosine monophosphate-activated protein kinase (AMPK) activation and its inhibition increases the efficacy of metformin in breast cancer cells. It also functions as a co-factor in the restriction imposed by tetherin on HIV-1, and targets HIV-1 Gag for lysosomal degradation, impairing virus assembly and release, in a tetherin-independent manner. Moreover, RNF115 is a Rab7-binding protein that stimulates c-Myc degradation through mono-ubiquitination of MM-1. It also plays crucial roles as a Rab7 target protein in vesicle traffic to late endosome/lysosome and lysosome biogenesis. Furthermore, RNF115 and the related protein, RNF126 associate with the epidermal growth factor receptor (EGFR) and promote ubiquitylation of EGFR, suggesting they play a role in the ubiquitin-dependent sorting and downregulation of membrane receptors. RNF115 contains an N-terminal BCA2 Zinc-finger domain (BZF), the AKT-phosphorylation sites, and the C-terminal C3H2C3-type RING-H2 finger. Pssm-ID: 438452 [Multi-domain] Cd Length: 50 Bit Score: 39.16 E-value: 3.81e-04
|
||||||||
| RING-HC_BRCA1 | cd16498 | RING finger, HC subclass, found in breast cancer type 1 susceptibility protein (BRCA1) and ... |
16-56 | 3.86e-04 | ||||
RING finger, HC subclass, found in breast cancer type 1 susceptibility protein (BRCA1) and similar proteins; BRCA1, also known as RING finger protein 53 (RNF53), is a RING finger protein encoded by the tumor suppressor gene BRCA1 that regulates all DNA double-strand break (DSB) repair pathways. BRCA1 is frequently mutated in patients with hereditary breast and ovarian cancer (HBOC). Its mutation is also associated with an increased risk of pancreatic, stomach, laryngeal, fallopian tube, and prostate cancer. It plays an important role in the DNA damage response signaling and has been implicated in various cellular processes such as cell cycle regulation, transcriptional regulation, chromatin remodeling, DNA DSBs, and apoptosis. BRCA1 contains an N-terminal C3HC4-type RING-HC finger, and two BRCT (BRCA1 C-terminus domain) repeats at the C-terminus. Pssm-ID: 438161 [Multi-domain] Cd Length: 94 Bit Score: 40.36 E-value: 3.86e-04
|
||||||||
| RING-HC_RNF169 | cd16551 | RING finger, HC subclass, found in RING finger protein 169 (RNF169) and similar proteins; ... |
15-60 | 3.86e-04 | ||||
RING finger, HC subclass, found in RING finger protein 169 (RNF169) and similar proteins; RNF169 is an uncharacterized E3 ubiquitin-protein ligase paralogous to RNF168. It functions as a negative regulator of the DNA damage signaling cascade. RNF169 recognizes polyubiquitin structures but does not itself contribute to double-strand break (DSB)-induced chromatin ubiquitylation. It contributes to regulation of the DSB repair pathway utilization via functionally competing with recruiting repair factors, 53BP1 and RAP80-BRCA1, for association with RNF168-modified chromatin independent of its catalytic activity, limiting the magnitude of the RNF8/RNF168-dependent signaling response to DSBs. RNF169 contains an N-terminal C3HC4-type RING-HC finger and a C-terminal MIU (motif interacting with ubiquitin) domain. Pssm-ID: 438213 [Multi-domain] Cd Length: 55 Bit Score: 39.45 E-value: 3.86e-04
|
||||||||
| RING-HC_AtRMA-like | cd16745 | RING finger, HC subclass, found in Arabidopsis thaliana RING membrane-anchor proteins (AtRMAs) ... |
18-53 | 3.98e-04 | ||||
RING finger, HC subclass, found in Arabidopsis thaliana RING membrane-anchor proteins (AtRMAs) and similar proteins; AtRMAs, including AtRma1, AtRma2, and AtRma3, are endoplasmic reticulum (ER)-localized Arabidopsis homologs of human outer membrane of the ER-anchor E3 ubiquitin-protein ligase, RING finger protein 5 (RNF5). AtRMAs possess E3 ubiquitin ligase activity, and may play a role in the growth and development of Arabidopsis. The AtRMA1 and AtRMA3 genes are predominantly expressed in major tissues, such as cotyledons, leaves, shoot-root junction, roots, and anthers, while AtRMA2 expression is restricted to the root tips and leaf hydathodes. AtRma1 probably functions with the Ubc4/5 subfamily of E2. AtRma2 is likely involved in the cellular regulation of ABP1 expression levels through interacting with auxin binding protein 1 (ABP1). AtRMA proteins contain an N-terminal C3HC4-type RING-HC finger and a trans-membrane-anchoring domain in their extreme C-terminal region. Pssm-ID: 438403 [Multi-domain] Cd Length: 45 Bit Score: 39.00 E-value: 3.98e-04
|
||||||||
| PDZ_TAX1BP3-like | cd10822 | PDZ domain of tax1-binding protein 3 (TAX1BP3), and related domains; PDZ (PSD-95 (Postsynaptic ... |
442-502 | 4.09e-04 | ||||
PDZ domain of tax1-binding protein 3 (TAX1BP3), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of TAX1BP3, and related domains. TAX1BP3 (also known as glutaminase-interacting protein 3, tax interaction protein 1, TIP-1, tax-interacting protein 1) may regulate a number of protein-protein interactions by competing for PDZ domain binding sites. TAX1BP3 binds beta-catenin and may act as an inhibitor of the Wnt signaling pathway. It competes with LIN7A (also known as Lin-7A or LIN-7A) for inward rectifier potassium channel 4 (KCNJ4) binding, and thereby promotes KCNJ4 internalization. It may play a role in the Rho signaling pathway, and in the activation of CDC42 by the viral protein HPV16 E6. Binding partners of the TAX1BP3 PDZ domain include beta-catenin, KCNJ4, glutaminase liver isoform (GLS2), rho guanine nucleotide exchange factor 16 (ARHGEF16), rhotekin, and CDK5 regulatory subunit-associated protein 3 (also known as LAPZ). PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This TAX1BP3-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467265 [Multi-domain] Cd Length: 94 Bit Score: 40.40 E-value: 4.09e-04
|
||||||||
| RING-HC_RNF5-like | cd16534 | RING finger, HC subclass, found in RING finger protein RNF5, RNF185 and similar proteins; RNF5 ... |
18-53 | 4.41e-04 | ||||
RING finger, HC subclass, found in RING finger protein RNF5, RNF185 and similar proteins; RNF5 and RNF185 are E3 ubiquitin-protein ligases that are anchored to the outer membrane of the endoplasmic reticulum (ER). RNF5 acts at early stages of cystic fibrosis (CF) transmembrane conductance regulator (CFTR) biosynthesis, and functions as a target for therapeutic modalities to antagonize mutant CFTR proteins in CF patients carrying the F508del allele. RNF185 controls the degradation of CFTR and CFTR F508del allele in a RING- and proteasome-dependent manner, but does not control that of other classical endoplasmic reticulum-associated degradation (ERAD) model substrates. Moreover, both RNF5 and RNF185 play important roles in cell adhesion and migration through the modulation of cell migration by ubiquitinating paxillin. Arabidopsis thaliana RING membrane-anchor proteins (AtRMAs) are also included in this family. They possess E3 ubiquitin-protein ligase activity and may play a role in the growth and development of Arabidopsis. All members of this family contain a C3HC4-type RING-HC finger. Pssm-ID: 438196 [Multi-domain] Cd Length: 44 Bit Score: 38.82 E-value: 4.41e-04
|
||||||||
| PDZ_MPP5-like | cd06798 | PDZ domain of membrane palmitoylated protein 5 (MPP5), Drosophila Stardust, and related ... |
427-478 | 4.44e-04 | ||||
PDZ domain of membrane palmitoylated protein 5 (MPP5), Drosophila Stardust, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of MPP5, Drosophila Stardust, and related domains. MPP5 (also known as MAGUK p55 subfamily member 1, protein associated with Lin-7 1 or PALS1) and Drosophila Stardust are membrane-associated guanylate kinase (MAGUK)-like proteins that serve as signaling and scaffolding proteins, linking different proteins critical to the formation and maintenance of tight junctions (TJ) and apical-basal polarity. Apical-basal polarity determinants cluster in complexes; in particular, the Crumbs complex (Crb, MPP5, and PATJ) and the PAR/aPKC-complex (PAR-3, PAR-6, aPKC) determine the apical plasma membrane domain. Within the Crumbs complex, Crb is stabilized in the plasma membrane by MPP5, which in turn recruits PATJ and Lin-7 to the complex. MPP5 also links the Crumbs complex with the PAR/aPKC-complex. The Drosophila homolog of the Crumbs complex is the (CRB)-Stardust (Sdt)-Discs Lost (Dlt) complex. MPP5 also acts as an interaction partner for SARS-CoV envelope protein E, which results in delayed formation of TJs and dysregulation of cell polarity. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MPP5-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467259 [Multi-domain] Cd Length: 79 Bit Score: 40.02 E-value: 4.44e-04
|
||||||||
| RING-HC_BAR | cd16497 | RING finger, HC subclass, found in bifunctional apoptosis regulator (BAR); BAR, also known as ... |
15-57 | 4.47e-04 | ||||
RING finger, HC subclass, found in bifunctional apoptosis regulator (BAR); BAR, also known as RING finger protein 47, was originally identified as an inhibitor of Bax-induced apoptosis. It participates in the block of apoptosis induced by TNF-family death receptors (extrinsic pathway) and mitochondria-dependent apoptosis (intrinsic pathway). BAR is predominantly expressed by neurons in the central nervous system and is involved in the regulation of neuronal survival. It is an endoplasmic reticulum (ER)-associated RING-type E3 ubiquitin ligase that interacts with BI-1 protein and post-translationally regulates its stability, as well as functioning in ER stress. BAR contains an N-terminal C3HC4-type RING-HC finger, a SAM domain, a coiled-coil domain, and a C-terminal transmembrane (TM) domain. This model corresponds to the RING-HC finger responsible for the binding of ubiquitin conjugating enzymes (E2s). Pssm-ID: 438160 [Multi-domain] Cd Length: 52 Bit Score: 39.03 E-value: 4.47e-04
|
||||||||
| RING-H2_RNF126 | cd16801 | RING finger, H2 subclass, found in RING finger protein 126 (RNF126) and similar proteins; ... |
17-57 | 4.51e-04 | ||||
RING finger, H2 subclass, found in RING finger protein 126 (RNF126) and similar proteins; RNF126 is a Bag6-dependent E3 ubiquitin ligase that is involved in the mislocalized protein (MLP) pathway of quality control. It regulates the retrograde sorting of the cation-independent mannose 6-phosphate receptor (CI-MPR). Moreover, RNF126 promotes cancer cell proliferation by targeting the tumor suppressor p21 for ubiquitin-mediated degradation, and could be a novel therapeutic target in breast and prostate cancers. It is also able to ubiquitylate cytidine deaminase (AID), a poorly soluble protein that is essential for antibody diversification. In addition, RNF126 and the related protein, RNF115 associate with the epidermal growth factor receptor (EGFR) and promote ubiquitylation of EGFR, suggesting they play a role in the ubiquitin-dependent sorting and downregulation of membrane receptors. RNF126 contains an N-terminal BCA2 Zinc-finger domain (BZF), the AKT-phosphorylation sites, and the C-terminal C3H2C3-type RING-H2 finger. Pssm-ID: 438453 [Multi-domain] Cd Length: 44 Bit Score: 38.82 E-value: 4.51e-04
|
||||||||
| RING-HC_TRIM58_C-IV | cd16606 | RING finger, HC subclass, found in tripartite motif-containing protein TRIM58 and similar ... |
15-58 | 4.86e-04 | ||||
RING finger, HC subclass, found in tripartite motif-containing protein TRIM58 and similar proteins; TRIM58, also known as protein BIA2, is an erythroid E3 ubiquitin-protein ligase induced during late erythropoiesis. It binds and ubiquitinates the intermediate chain of the microtubule motor dynein (DYNC1LI1/DYNC1LI2), stimulating the degradation of the dynein holoprotein complex. It may participate in the erythroblast enucleation process through regulation of nuclear polarization. TRIM58 belongs to the C-IV subclass of TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region, as well as a B30.2/SPRY (SplA and ryanodine receptor) domain positioned C-terminal to the RBCC domain. Pssm-ID: 438268 [Multi-domain] Cd Length: 53 Bit Score: 39.07 E-value: 4.86e-04
|
||||||||
| PDZ5_INAD-like | cd23066 | PDZ domain 5 of inactivation no after potential D (INAD), and related domains; PDZ (PSD-95 ... |
445-502 | 4.87e-04 | ||||
PDZ domain 5 of inactivation no after potential D (INAD), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 5 of INAD, and related domains. INAD assembles key enzymes of the Drosophila compound eye photo-transduction pathway into a supramolecular complex, supporting efficient and fast light signaling. It contains 5 PDZ domains arranged in tandem (PDZ1-PDZ5) which independently bind various proteins. INAD PDZ2 binds eye-specific protein kinase C, INAD PDZ3 binds transient receptor potential (TRP) channel, and INAD PDZ45 tandem binds NORPA (phospholipase Cbeta, PLCbeta). Mutations of the inaD gene that lead to disruption of each of these interactions impair fly photo signal transduction. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This INAD-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467279 [Multi-domain] Cd Length: 80 Bit Score: 39.79 E-value: 4.87e-04
|
||||||||
| RING-HC_RNF146 | cd16546 | RING finger, HC subclass, found in RING finger protein 146 (RNF146) and similar proteins; ... |
16-57 | 4.87e-04 | ||||
RING finger, HC subclass, found in RING finger protein 146 (RNF146) and similar proteins; RNF146, also known as dactylidin, or iduna, is a cytoplasmic E3 ubiquitin-protein ligase that is responsible for PARylation-dependent ubiquitination (PARdU). It displays neuroprotective property due to its inhibition of Parthanatos, a PAR dependent cell death, via binding with Poly(ADP-ribose) (PAR). It also modulates PAR polymerase-1 (PARP-1)-mediated oxidative cell injury in cardiac myocytes. Moreover, RNF146 mediates tankyrase-dependent degradation of axin, thereby positively regulating Wnt signaling. It also facilitates DNA repair and protects against cell death induced by DNA damaging agents or gamma-irradiation by translocating to the nucleus after cellular injury and promoting the ubiquitination and degradation of various nuclear proteins involved in DNA damage repair. Furthermore, RNF146 is implicated in neurodegenerative disease and cancer development. It regulates the development and progression of non-small cell lung cancer (NSCLC) by enhancing cell growth, invasion, and survival. RNF146 contains an N-terminal C3HC4-type RING-HC finger followed by a WWE domain with a poly(ADP-ribose) (PAR) binding motif at the tail. Pssm-ID: 438208 [Multi-domain] Cd Length: 50 Bit Score: 38.91 E-value: 4.87e-04
|
||||||||
| PDZ5_MUPP1-like | cd06669 | PDZ domain 5 of multi-PDZ-domain protein 1 (MUPP1), PATJ (protein-associated tight junction) ... |
441-504 | 4.99e-04 | ||||
PDZ domain 5 of multi-PDZ-domain protein 1 (MUPP1), PATJ (protein-associated tight junction) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 5 of MUPP1 and PATJ, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ5 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F Pssm-ID: 467157 [Multi-domain] Cd Length: 98 Bit Score: 40.29 E-value: 4.99e-04
|
||||||||
| cpPDZ_CPP-like | cd06782 | circularly permuted PDZ domain of C-terminal processing peptidase (CPP), a serine protease, ... |
277-337 | 5.04e-04 | ||||
circularly permuted PDZ domain of C-terminal processing peptidase (CPP), a serine protease, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of CPP (also known as tail-specific protease, PRC protein, Protease Re, and Photosystem II D1 protein processing peptidase), and related domains. CPP belongs to the peptidase S41A family. It cleaves a C-terminal 11 residue peptide from the precursor form of penicillin-binding protein 3, and may have a role in protecting bacterium from thermal and osmotic stresses. In the plant chloroplast, the enzyme removes the C-terminal extension of the D1 polypeptide of photosystem II. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains and as well as those with circular permutations and domain swapping of beta-strands. The canonical PDZ domain contains six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. This CPP-like PDZ domain is a circularly permuted PDZ domain which places beta-strand A on the C-terminus. Another permutation exists in the PDZ superfamily which places both beta-strands A and B on the C-terminus. Pssm-ID: 467623 [Multi-domain] Cd Length: 88 Bit Score: 40.16 E-value: 5.04e-04
|
||||||||
| PDZ_PICK1-like | cd06722 | PDZ domain of PICK1 (protein interacting with C-kinase 1) and similar domains; PDZ (PSD-95 ... |
434-481 | 5.43e-04 | ||||
PDZ domain of PICK1 (protein interacting with C-kinase 1) and similar domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of PICK1, and related domains. PICK1 (also known as PRKCA-binding protein and protein kinase C-alpha-binding protein) plays a key role in regulating trafficking of binding partners by altering either their subcellular targeting and/or surface expression. PICK1 plays a role in synaptic plasticity by regulating the trafficking and internalization of amino-3-hydroxy-5-methylisoxazole-4-propionic acid (AMPA) receptors; the PICK1-PDZ domain binds the AMPA receptor subunits. The PICK1 PDZ domain also binds glutamate transporters, Eph receptors, metabotropic glutamate receptors, and ASICs (acid-sensing ion channels), among others. Clustering and synaptic targeting of PICK1 requires direct interaction between the PDZ domain and lipid membranes. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This PICK-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged as beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta- strand F. Pssm-ID: 467205 [Multi-domain] Cd Length: 84 Bit Score: 39.71 E-value: 5.43e-04
|
||||||||
| NifB | COG1625 | Fe-S oxidoreductase, related to NifB/MoaA family [Energy production and conversion]; |
446-478 | 6.24e-04 | ||||
Fe-S oxidoreductase, related to NifB/MoaA family [Energy production and conversion]; Pssm-ID: 441232 [Multi-domain] Cd Length: 441 Bit Score: 43.59 E-value: 6.24e-04
|
||||||||
| SdrC | COG3480 | Predicted secreted protein YlbL, contains PDZ domain [Signal transduction mechanisms]; |
278-337 | 6.52e-04 | ||||
Predicted secreted protein YlbL, contains PDZ domain [Signal transduction mechanisms]; Pssm-ID: 442703 [Multi-domain] Cd Length: 344 Bit Score: 43.26 E-value: 6.52e-04
|
||||||||
| cpPDZ_DegS | cd06777 | circularly permuted PDZ domain of DegS serine endoprotease; PDZ (PSD-95 (Postsynaptic density ... |
259-337 | 6.98e-04 | ||||
circularly permuted PDZ domain of DegS serine endoprotease; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of Escherichia coli DegS and related domains. DegS (also known as Site-1 protease DegS, S1P protease DegS, and Site-1-type intramembrane protease) participates in the activation of the sigma(E) extracytoplasmic stress response. Initially, there is an accumulation of misfolded membrane proteins (OMPs) in the periplasm which bind by their YXF motif to the DegS PDZ domain, activating DegS-catalyzed cleavage of the RseA periplasmic domain and making RseA a substrate for cleavage by another membrane protease RseP. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains and as well as those with circular permutations and domain swapping of beta-strands. The canonical PDZ domain contains six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. This DegS family PDZ domain is a circularly permuted PDZ domain which places beta-strand A on the C-terminus. Another permutation exists in the PDZ superfamily which places both beta-strands A and B on the C-terminus. Pssm-ID: 467620 [Multi-domain] Cd Length: 93 Bit Score: 39.68 E-value: 6.98e-04
|
||||||||
| RING-HC_TRIM5-like_C-IV | cd16591 | RING finger, HC subclass, found in tripartite motif-containing proteins TRIM5, TRIM6, TRIM22, ... |
10-65 | 6.99e-04 | ||||
RING finger, HC subclass, found in tripartite motif-containing proteins TRIM5, TRIM6, TRIM22, TRIM34 and similar proteins; TRIM5, TRIM6, TRIM22, and TRIM34, four closely related tripartite motif-containing proteins, belong to the C-IV subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region, as well as a B30.2/SPRY (SplA and ryanodine receptor) domain positioned C-terminal to the RBCC domain. TRIM5, also known as RING finger protein 88 (RNF88), is a capsid-specific restriction factor that prevents infection from non-host-adapted retroviruses in a species-specific manner by binding to and destabilizing the retroviral capsid lattice before reverse transcription is completed. Its retroviral restriction activity correlates with the ability to activate TAK1-dependent innate immune signaling. TRIM5 also acts as a pattern recognition receptor that activates innate immune signaling in response to the retroviral capsid lattice. Moreover, TRIM5 plays a role in regulating autophagy through activation of autophagy regulator BECN1 by causing its dissociation from its inhibitors BCL2 and TAB2. It also plays a role in autophagy by acting as a selective autophagy receptor which recognizes and targets HIV-1 capsid protein p24 for autophagic destruction. TRIM6, also known as RING finger protein 89 (RNF89), is an E3-ubiquitin ligase that cooperates with the E2-ubiquitin conjugase UbE2K to catalyze the synthesis of unanchored K48-linked polyubiquitin chains, and further stimulates the interferon-I kappa B kinase epsilon (IKKepsilon) kinase-mediated antiviral response. It also regulates the transcriptional activity of Myc during the maintenance of embryonic stem (ES) cell pluripotency, and may act as a novel regulator for Myc-mediated transcription in ES cells. TRIM22, also known as 50 kDa-stimulated trans-acting factor (Staf-50) or RING finger protein 94 (RNF94), is an E3 ubiquitin-protein ligase that plays an integral role in the host innate immune response to viruses. It has been shown to inhibit the replication of a number of viruses, including HIV-1, hepatitis B, and influenza A. TRIM22 acts as a suppressor of basal HIV-1 long terminal repeat (LTR)-driven transcription by preventing the transcription factor specificity protein 1 (Sp1) binding to the HIV-1 promoter. It also controls FoxO4 activity and cell survival by directing Toll-like receptor 3 (TLR3)-stimulated cells toward type I interferon (IFN) type I gene induction or apoptosis. Moreover, TRIM22 can activate the noncanonical nuclear factor-kappaB (NF-kappaB) pathway by activating I kappa B kinase alpha (IKKalpha). It also regulates nucleotide binding oligomerization domain containing 2 (NOD2)-dependent activation of interferon-beta signaling and nuclear factor-kappaB. TRIM34, also known as interferon-responsive finger protein 1 or RING finger protein 21 (RNF21), may function as antiviral protein that contribute to the defense against retroviral infections. Pssm-ID: 438253 [Multi-domain] Cd Length: 72 Bit Score: 39.35 E-value: 6.99e-04
|
||||||||
| RING-HC_TRIM69_C-IV | cd16611 | RING finger, HC subclass, found in tripartite motif-containing protein 69 (TRIM69) and similar ... |
15-57 | 7.40e-04 | ||||
RING finger, HC subclass, found in tripartite motif-containing protein 69 (TRIM69) and similar proteins; TRIM69, also known as RFP-like domain-containing protein trimless or RING finger protein 36 (RNF36), is a testis E3 ubiquitin-protein ligase that plays a specific role in apoptosis and may also play an important role in germ cell homeostasis during spermatogenesis. TRIM69 belongs to the C-IV subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region, as well as a B30.2/SPRY (SplA and ryanodine receptor) domain positioned C-terminal to the RBCC domain. Pssm-ID: 438273 [Multi-domain] Cd Length: 59 Bit Score: 38.59 E-value: 7.40e-04
|
||||||||
| RING-HC_RNF8 | cd16535 | RING finger, HC subclass, found in RING finger protein 8 (RNF8) and similar proteins; RNF8 is ... |
15-69 | 7.54e-04 | ||||
RING finger, HC subclass, found in RING finger protein 8 (RNF8) and similar proteins; RNF8 is a telomere-associated E3 ubiquitin-protein ligase that plays an important role in DNA double-strand break (DSB) repair via histone ubiquitination. It is localized in the nucleus and interacts with class III E2s (UBE2E2, UbcH6, and UBE2E3), but not with other E2s (UbcH5, UbcH7, UbcH10, hCdc34, and hBendless). It recruits UBC13 for lysine 63-based self polyubiquitylation. Its deficiency causes neuronal pathology and cognitive decline, and its loss results in neuron degeneration. RNF8, together with RNF168, catalyzes a series of ubiquitylation events on substrates such as H2A and H2AX, with the H2AK13/15 ubiquitylation being particularly important for recruitment of repair factors p53-binding protein 1 (53BP1) or the RAP80-BRCA1 complex to sites of DSBs. RNF8 mediates the ubiquitination of gammaH2AX, and recruits 53BP1 and BRCA1 to DNA damage sites which promotes DNA damage response (DDR) and inhibits chromosomal instability. Moreover, RNF8 interacts with retinoid X receptor alpha (RXR alpha) and enhances its transcription-stimulating activity. It also regulates the rate of exit from mitosis and cytokinesis. RNF8 contains an N-terminal forkhead-associated (FHA) domain and a C-terminal C3HC4-type RING-HC finger. Pssm-ID: 438197 [Multi-domain] Cd Length: 64 Bit Score: 38.91 E-value: 7.54e-04
|
||||||||
| RING-HC_TRIM9 | cd16755 | RING finger, HC subclass, found in tripartite motif-containing protein 9 (TRIM9) and similar ... |
13-56 | 7.58e-04 | ||||
RING finger, HC subclass, found in tripartite motif-containing protein 9 (TRIM9) and similar proteins; TRIM9, human ortholog of rat Spring, also known as RING finger protein 91 (RNF91), is a brain-specific E3 ubiquitin-protein ligase collaborating with an E2 ubiquitin conjugating enzyme UBCH5b. TRIM9 plays an important role in the regulation of neuronal functions and participates in the neurodegenerative disorders through its ligase activity. It interacts with the WD repeat region of beta-transducin repeat-containing protein (beta-TrCP) through its N-terminal degron motif depending on the phosphorylation status, and thus negatively regulates nuclear factor-kappaB (NF-kappaB) activation in the NF-kappaB pro-inflammatory signaling pathway. Moreover, TRIM9 acts as a critical catalytic link between Netrin-1 and the exocytic soluble NSF attachment receptor protein (SNARE) machinery in murine cortical neurons. It promotes SNARE-mediated vesicle fusion and axon branching in a Netrin-dependent manner. TRIM9 belongs to the C-I subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region, as well as a COS (carboxyl-terminal subgroup one signature) box, a fibronectin type III (FN3) domain, and a B30.2/SPRY (SplA and ryanodine receptor) domain positioned C-terminal to the RBCC domain. Pssm-ID: 438413 [Multi-domain] Cd Length: 55 Bit Score: 38.47 E-value: 7.58e-04
|
||||||||
| RING-HC_TRIM41-like_C-IV | cd16602 | RING finger, HC subclass, found in tripartite motif-containing proteins TRIM41, TRIM52 and ... |
18-57 | 7.75e-04 | ||||
RING finger, HC subclass, found in tripartite motif-containing proteins TRIM41, TRIM52 and similar proteins; TRIM41 and TRIM52, two closely related tripartite motif-containing proteins, have dramatically expanded RING domains compared with the rest of the TRIM family proteins. TRIM41 belongs to the C-IV subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region, as well as a B30.2/SPRY (SplA and ryanodine receptor) domain positioned C-terminal to the RBCC domain. In contrast, TRIM52 lacks the putative viral recognition SPRY/B30.2 domain, and thus has been classified to the C-V subclass of the TRIM family that contains only RBCC domains. TRIM41, also known as RING finger-interacting protein with C kinase (RINCK), is an E3 ubiquitin-protein ligase that promotes the ubiquitination of protein kinase C (PKC) isozymes in cells. It specifically recognizes the C1 domain of PKC isozymes. It controls the amplitude of PKC signaling by controlling the amount of PKC in the cell. TRIM52, also known as RING finger protein 102 (RNF102), is encoded by a novel, noncanonical antiviral TRIM52 gene in primate genomes with unique specificity determined by the rapidly evolving RING domain. Pssm-ID: 438264 [Multi-domain] Cd Length: 53 Bit Score: 38.37 E-value: 7.75e-04
|
||||||||
| PDZ6_GRIP1-2-like | cd06683 | PDZ domain 6 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related ... |
249-337 | 8.81e-04 | ||||
PDZ domain 6 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR) binding proteins GRIP1 (ABP/GRIP2) and GRIP2, and related domains. GRIP1 and GRIP2 each have 7 PDZ domains. The interaction of GRIP1 and GRIP2 with GluA2/3 (AMPAR subunit) regulates AMPAR trafficking and synaptic targeting. GRIP1 has an essential role in regulating AMPAR trafficking during synaptic plasticity and learning and memory. GRIP1 and GRIP2 interact with a variety of other proteins associated with protein trafficking and internalization, for example GRIP1 also interacts with KIF5 (also known as kinesin 1), EphB receptors, scaffold protein liprin-alpha, and the rasGEF GRASP-1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This GRIP family PDZ6 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467171 [Multi-domain] Cd Length: 85 Bit Score: 39.21 E-value: 8.81e-04
|
||||||||
| RING-HC_TRIM26_C-IV | cd16598 | RING finger, HC subclass, found in tripartite motif-containing protein 26 (TRIM26) and similar ... |
15-42 | 9.32e-04 | ||||
RING finger, HC subclass, found in tripartite motif-containing protein 26 (TRIM26) and similar proteins; TRIM26, also known as acid finger protein (AFP), RING finger protein 95 (RNF95), or zinc finger protein 173 (ZNF173), is an E3 ubiquitin-protein ligase that negatively regulates interferon-beta production and antiviral response through polyubiquitination and degradation of nuclear transcription factor IRF3. It functions as an important regulator for RNA virus-triggered innate immune response by bridging TBK1 to NEMO (NF-kappaB essential modulator, also known as IKKgamma) and mediating TBK1 activation. It also acts as a novel tumor suppressor of hepatocellular carcinoma by regulating cancer cell proliferation, colony forming ability, migration, and invasion. TRIM26 belongs the C-IV subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, a B-box, and two coiled coil domains, as well as a B30.2/SPRY (SplA and ryanodine receptor) domain positioned C-terminal to the RBCC domain. Pssm-ID: 438260 [Multi-domain] Cd Length: 64 Bit Score: 38.61 E-value: 9.32e-04
|
||||||||
| PDZ_MPP3-MPP4-MPP7-like | cd06799 | PDZ domain of membrane palmitoylated proteins 3 (MPP3), MPP4, and MPP7, and related domains; ... |
425-489 | 1.04e-03 | ||||
PDZ domain of membrane palmitoylated proteins 3 (MPP3), MPP4, and MPP7, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of MPP3, MPP4, and MPP7, and related domains. MPP3 (also known as MAGUK p55 subfamily member 3, erythrocyte membrane protein p55, or EMP55), MPP4 (also known as MAGUK p55 subfamily member 4 or Discs large homolog 6), and MPP7 (also known as MAGUK p55 subfamily member 7) are membrane-associated guanylate kinase (MAGUK)-like proteins. MPP3 is part of a cell adhesion protein complex including tumor suppressor CADM1 and actin-binding protein 4.1B. Participation in the Crumbs cell polarity complex has also been demonstrated for MPP7 in epithelial cells, and for MPP3 and MPP4 in the retina. MPP4 is needed for proper localization of plasma membrane calcium ATPases and maintenance of calcium homeostasis at the rod photoreceptor synaptic terminals. Binding partners of the MPP3 PDZ domain include nectin-3, serotonin 5-hydroxytryptamine, 5-HT(2C) receptor, and a cell adhesion protein, TSLC1 (tumor suppressor in lung cancer 1); fragments of MPP4 having the PDZ domain bind CRB (PDZ-SH3-GUK) and GABA transporter GAT1 (PDZ-SH3). PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MPP1-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467260 [Multi-domain] Cd Length: 81 Bit Score: 38.76 E-value: 1.04e-03
|
||||||||
| RseP | COG0750 | Membrane-associated protease RseP, regulator of RpoE activity [Posttranslational modification, ... |
283-337 | 1.05e-03 | ||||
Membrane-associated protease RseP, regulator of RpoE activity [Posttranslational modification, protein turnover, chaperones, Transcription]; Pssm-ID: 440513 [Multi-domain] Cd Length: 349 Bit Score: 42.38 E-value: 1.05e-03
|
||||||||
| mRING-HC-C3HC3D_PHRF1 | cd16635 | Modified RING finger, HC subclass (C3HC3D-type), found in PHD and RING finger ... |
13-53 | 1.07e-03 | ||||
Modified RING finger, HC subclass (C3HC3D-type), found in PHD and RING finger domain-containing protein 1 (PHRF1) and similar proteins; PHRF1, also known as KIAA1542, or CTD-binding SR-like protein rA9, is a ubiquitin ligase which induces the ubiquitination of TGIF (TG-interacting factor) at lysine 130. It acts as a tumor suppressor that promotes the transforming growth factor (TGF)-beta cytostatic program through selective release of TGIF-driven promyelocytic leukemia protein (PML) inactivation. PHRF1 contains a plant homeodomain (PHD) finger and a modified C3HC3D-type RING-HC finger. Pssm-ID: 438297 [Multi-domain] Cd Length: 51 Bit Score: 38.17 E-value: 1.07e-03
|
||||||||
| PDZ_neurabin-like | cd06790 | PDZ domain of neurabin-1 and neurabin-2, and related domains; PDZ (PSD-95 (Postsynaptic ... |
442-476 | 1.08e-03 | ||||
PDZ domain of neurabin-1 and neurabin-2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of neurabin-1 (also known as protein phosphatase 1 regulatory subunit 9A) and neurabin-2 (also known as spinophilin, and protein phosphatase 1 regulatory subunit 9B), and related domains. Neurabin-1 and neurabin-2 are neuronal scaffolding proteins that play important roles in the regulation of synaptic transmission through their ability to interact with and target protein phosphatase 1 (PP1) to dendritic spines where PP1 dephosphorylates and inactivates glutamate receptors. Neurabin-2 interacts with multiple other synaptic proteins, including synaptic signaling and scaffolding proteins (e.g., GluN1 and SAPAP3) and cytoskeletal proteins (e.g., neurofilament medium polypeptide, NF-M). Neurabin-1 and neurabin-2 also binds F-actin. Other binding partners of neurabin-1 include adenosine A1 receptor (A1R), SAD-1 kinase and 70 kDa ribosomal protein S6 kinase (p70-S6K). This PDZ domain is immediately C-terminal to the PP1 binding domain. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This neurabin-like PDZ domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467252 [Multi-domain] Cd Length: 90 Bit Score: 39.32 E-value: 1.08e-03
|
||||||||
| PDZ_nNOS-like | cd06708 | PDZ domain of neuronal nitric oxide synthase (nNOS), and related domains; PDZ (PSD-95 ... |
281-337 | 1.19e-03 | ||||
PDZ domain of neuronal nitric oxide synthase (nNOS), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of nNOS, and related domains. nNOS produces a key signaling molecule, nitric oxide (NO), which has diverse functions throughout the body and acts as a neurotransmitter and intracellular signaling molecule in the central and peripheral nervous system. nNOS is concentrated at synaptic junctions in the brain and motor endplates in skeletal muscle. The PDZ domain of neuronal nitric oxide synthase (nNOS) interacts with the PDZ domain of alpha1-syntrophin (in muscle cells) and with the second PDZ domain of Disks large homolog 4 (Dlg4, also known as PSD-95), and nitric oxide synthase 1 adaptor protein NOS1AP in neurons. Dlg4 binds NMDA receptors, and nNOS, forming a complex in neurons. NOS1AP competes with Dgl4 for the nNOS PDZ domain and prevents the coupling of nNos activation with NMDA receptor-mediated calcium influx. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This nNOS-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467192 [Multi-domain] Cd Length: 110 Bit Score: 39.67 E-value: 1.19e-03
|
||||||||
| PDZ_MPP-like | cd06726 | PDZ domain of membrane palmitoylated proteins (MPPs), and related domains; PDZ (PSD-95 ... |
431-500 | 1.20e-03 | ||||
PDZ domain of membrane palmitoylated proteins (MPPs), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of MPP1-7 (also known as MAGUK p55 subfamily members 1-7), and related domains. MPPs comprise a subfamily of a larger group of multidomain proteins, namely, membrane-associated guanylate kinases (MAGUKs). MPPs form diverse protein complexes at the cell membranes, which are involved in a wide range of cellular processes, including establishing proper cell structure, polarity and cell adhesion. MPPs have only one PDZ domain. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MPP1-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467208 [Multi-domain] Cd Length: 80 Bit Score: 38.79 E-value: 1.20e-03
|
||||||||
| cpPDZ_HtrA-like | cd06785 | circularly permuted PDZ domain of high-temperature requirement factor A (HtrA) family serine ... |
438-493 | 1.34e-03 | ||||
circularly permuted PDZ domain of high-temperature requirement factor A (HtrA) family serine proteases and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of HtrA family serine proteases including human HtrA1, HtrA2 (mitochondrial), HtrA3, and HtrA4, and related domains. These proteases are key enzymes associated with pregnancy. Their diverse biological functions include cell growth proliferation, migration and apoptosis. They are also implicated in disorders including Alzheimer's, Parkinson's, arthritis and cancer. HtrA1 (also known as high-temperature requirement A serine peptidase 1, L56, and serine protease 11) substrates include extracellular matrix proteins, proteoglycans, and insulin-like growth factor (IGF)-binding proteins. HtrA1 also inhibits signaling by members of the transforming growth factor beta (TGF-beta) family. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping of beta-strands. The canonical PDZ domain contains six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. This HtrA-like PDZ domain is a circularly permuted PDZ domain which places beta-strand A on the C-terminus. Another permutation exists in the PDZ superfamily which places both beta-strands A and B on the C-terminus. Pssm-ID: 467624 [Multi-domain] Cd Length: 98 Bit Score: 39.02 E-value: 1.34e-03
|
||||||||
| RING-HC_UHRF | cd16613 | RING finger, HC subclass, found in ubiquitin-like PHD and RING finger domain-containing ... |
17-60 | 1.35e-03 | ||||
RING finger, HC subclass, found in ubiquitin-like PHD and RING finger domain-containing proteins, UHRF1 and UHRF2, and similar proteins; UHRF1 is a unique chromatin effector protein that integrates the recognition of both histone PTMs and DNA methylation. It is essential for cell proliferation and plays a critical role in the development and progression of many human carcinomas, such as laryngeal squamous cell carcinoma (LSCC), gastric cancer (GC), esophageal squamous cell carcinoma (ESCC), colorectal cancer, prostate cancer, and breast cancer. UHRF1 acts as a transcriptional repressor through its binding to histone H3 when it is unmodified at Arg2. Its overexpression in human lung fibroblasts results in downregulation of expression of the tumor suppressor pRB. It also plays a role in transcriptional repression of the cell cycle regulator p21. Moreover, UHRF1-dependent repression of transcription factors can facilitate the G1-S transition. It interacts with Tat-interacting protein of 60 kDa (TIP60) and induces degradation-independent ubiquitination of TIP60. It is also an N-methylpurine DNA glycosylase (MPG)-interacting protein that binds MPG in a p53 status-independent manner in the DNA base excision repair (BER) pathway. In addition, UHRF1 functions as an epigenetic regulator that is important for multiple aspects of epigenetic regulation, including maintenance of DNA methylation patterns and recognition of various histone modifications. UHRF2 was originally identified as a ubiquitin ligase acting as a small ubiquitin-like modifier (SUMO) E3 ligase that enhances zinc finger protein 131 (ZNF131) SUMOylation, but does not enhance ZNF131 ubiquitination. It also ubiquitinates PCNP, a PEST-containing nuclear protein. Moreover, UHRF2 functions as a nuclear protein involved in cell-cycle regulation and has been implicated in tumorigenesis. It interacts with cyclins, CDKs, p53, pRB, PCNA, HDAC1, DNMTs, G9a, methylated histone H3 lysine 9, and methylated DNA. It interacts with the cyclin E-CDK2 complex, ubiquitinates cyclins D1 and E1, induces G1 arrest, and is involved in the G1/S transition regulation. Furthermore, UHRF2 is a direct transcriptional target of the transcription factor E2F-1 in the induction of apoptosis. It recruits HDAC1 and binds to methyl-CpG. UHRF2 also participates in the maturation of Hepatitis B virus (HBV) by interacting with the HBV core protein and promoting its degradation. Both UHRF1 and UHRF2 contain an N-terminal ubiquitin-like domain (UBL), a tandem Tudor domain (TTD), a plant homeodomain (PHD) finger, a SET- and RING-associated (SRA) domain, and a C-terminal C3HC4-type RING-HC finger. Pssm-ID: 438275 [Multi-domain] Cd Length: 46 Bit Score: 37.72 E-value: 1.35e-03
|
||||||||
| RING-HC_TRIM9-like_C-I | cd16576 | RING finger, HC subclass, found in tripartite motif-containing proteins TRIM9, TRIM67, and ... |
13-57 | 1.38e-03 | ||||
RING finger, HC subclass, found in tripartite motif-containing proteins TRIM9, TRIM67, and similar proteins; Tripartite motif-containing proteins TRIM9 and TRIM67 belong to the C-I subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, consisting of three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region, as well as a COS (carboxyl-terminal subgroup one signature) box, a fibronectin type III (FN3) domain, and a B30.2/SPRY (SplA and ryanodine receptor) domain positioned C-terminal to the RBCC domain. TRIM9 (the human ortholog of rat Spring), also known as RING finger protein 91 (RNF91), is a brain-specific E3 ubiquitin-protein ligase collaborating with an E2 ubiquitin conjugating enzyme UBCH5b. TRIM9 plays an important role in the regulation of neuronal functions and participates in neurodegenerative disorders through its ligase activity. TRIM67, also known as TRIM9-like protein (TNL), is a protein selectively expressed in the cerebellum. It interacts with PRG-1, an important molecule in the control of hippocampal excitability dependent on presynaptic LPA2 receptor signaling, and 80K-H, also known as glucosidase II beta, a protein kinase C substrate. Pssm-ID: 438238 [Multi-domain] Cd Length: 42 Bit Score: 37.39 E-value: 1.38e-03
|
||||||||
| RING-HC_TRIM13_like_C-V | cd16581 | RING finger, HC subclass, found in tripartite motif-containing proteins TRIM13, TRIM59 and ... |
15-42 | 1.41e-03 | ||||
RING finger, HC subclass, found in tripartite motif-containing proteins TRIM13, TRIM59 and similar proteins; TRIM13 and TRIM59, two closely related tripartite motif-containing proteins, belong to the C-V subclass of the TRIM (tripartite motif) family of proteins that are defined by an N-terminal RBCC (RING, Bbox, and coiled coil) domain, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region, followed by a C-terminal transmembrane domain. TRIM13, also known as B-cell chronic lymphocytic leukemia tumor suppressor Leu5, leukemia-associated protein 5, putative tumor suppressor RFP2, RING finger protein 77 (RNF77), or Ret finger protein 2, is an endoplasmic reticulum (ER) membrane anchored E3 ubiquitin-protein ligase that interacts with proteins localized to the ER, including valosin-containing protein (VCP), a protein indispensable for ER-associated degradation (ERAD). TRIM59, also known as RING finger protein 104 (RNF104) or tumor suppressor TSBF-1, is a putative E3 ubiquitin-protein ligase that functions as a novel multiple cancer biomarker for immunohistochemical detection of early tumorigenesis. Pssm-ID: 438243 [Multi-domain] Cd Length: 50 Bit Score: 37.49 E-value: 1.41e-03
|
||||||||
| RING-HC_BAH1-like | cd23127 | RING finger, HC subclass, found in Arabidopsis thaliana protein BENZOIC ACID HYPERSENSITIVE 1 ... |
12-41 | 1.49e-03 | ||||
RING finger, HC subclass, found in Arabidopsis thaliana protein BENZOIC ACID HYPERSENSITIVE 1 (BAH1) and similar proteins; This subfamily includes Arabidopsis thaliana BAH1 and BAH1-like. BAH1, also known as protein NITROGEN LIMITATION ADAPTATION (NLA), or RING-type E3 ubiquitin transferase BAH1, acts as an E3 ubiquitin-protein ligase that mediates E2-dependent protein ubiquitination. It plays a role in salicylic acid-mediated negative feedback regulation of salicylic acid (SA) accumulation. It may be involved in the overall regulation of SA, benzoic acid and phenylpropanoid biosynthesis. It controls the adaptability to nitrogen limitation by channeling the phenylpropanoid metabolic flux to the induced anthocyanin synthesis. BAH1-like, also known as RING finger protein 178, or RING-type E3 ubiquitin transferase BAH1-like, is a probable E3 ubiquitin-protein ligase. Members of this subfamily contain a typical C3HC4-type RING-HC finger. Pssm-ID: 438489 [Multi-domain] Cd Length: 74 Bit Score: 38.15 E-value: 1.49e-03
|
||||||||
| PDZ6_MUPP1-like | cd06670 | PDZ domain 6 of multi-PDZ-domain protein 1 (MUPP1) and related domains; PDZ (PSD-95 ... |
431-475 | 1.55e-03 | ||||
PDZ domain 6 of multi-PDZ-domain protein 1 (MUPP1) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 6 of multi-PDZ-domain protein 1 (MUPP1). MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, PDZ9, and PDZ13. This MuPP1-like PDZ6 domain is therefore absent from PATJ. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ6 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F Pssm-ID: 467158 [Multi-domain] Cd Length: 87 Bit Score: 38.77 E-value: 1.55e-03
|
||||||||
| PDZ4_DLG5-like | cd06766 | PDZ domain 4 of Discs Large 5 (Dlg5) and related domains; PDZ (PSD-95 (Postsynaptic density ... |
427-493 | 1.56e-03 | ||||
PDZ domain 4 of Discs Large 5 (Dlg5) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 4 of Drosophila and mammalian Dlg5, and related domains. Dlg5 is a scaffold protein with multiple conserved functions that are independent of each other in regulating growth, cell polarity, and cell adhesion. It has a coiled-coil domain, 4 PDZ domains and a MAGUK domain (an SH3 domain next to a non-catalytically active guanylate kinase domain). Deregulation of Dlg5 has been implicated in the malignancy of several cancer types. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Dlg5-like family PDZ4 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467247 [Multi-domain] Cd Length: 81 Bit Score: 38.52 E-value: 1.56e-03
|
||||||||
| PDZ_GOPC-like | cd06800 | PDZ domain of Golgi-associated PDZ and coiled-coil motif-containing protein (GOPC), and ... |
442-492 | 1.59e-03 | ||||
PDZ domain of Golgi-associated PDZ and coiled-coil motif-containing protein (GOPC), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of GOPC and related domains. GOPC, also known as PIST (PDZ domain protein interacting specifically with TC10), FIG (fused in glioblastoma), and CAL (CFTR-associated ligand), regulates the trafficking of a wide array of proteins, including small GTPases, receptors, and cell surface molecules such as cadherin 23 and CFTR. It may regulate CFTR chloride currents and acid-sensing ASIC3 currents by modulating cell surface expression of both channels, and may play a role in autophagy. Interaction partners of the GOPC PDZ domains include: FZD5, FZD8, ASIC3, CFTR, MUC3, ARFRP1, Ggamma13, neuroligin, and Stargazin. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This GOPC-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467261 [Multi-domain] Cd Length: 83 Bit Score: 38.51 E-value: 1.59e-03
|
||||||||
| PDZ7_GRIP1-2-like | cd06685 | PDZ domain 7 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related ... |
420-502 | 1.62e-03 | ||||
PDZ domain 7 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR) binding proteins GRIP1 (ABP/GRIP2) and GRIP2, and related domains. GRIP1 and GRIP2 each have 7 PDZ domains. The interaction of GRIP1 and GRIP2 with GluA2/3 (AMPAR subunit) regulates AMPAR trafficking and synaptic targeting. GRIP1 has an essential role in regulating AMPAR trafficking during synaptic plasticity and learning and memory. GRIP1 and GRIP2 interact with a variety of other proteins associated with protein trafficking and internalization, for example GRIP1 also interacts with KIF5 (also known as kinesin 1), EphB receptors, scaffold protein liprin-alpha, and the rasGEF GRASP-1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This GRIP family PDZ7 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467173 [Multi-domain] Cd Length: 85 Bit Score: 38.39 E-value: 1.62e-03
|
||||||||
| PDZ_CNK1_2_3-like | cd06748 | PDZ domain of connector enhancer of kinase suppressor of ras 1 (CNK1), CNK2, CNK3, and related ... |
438-475 | 1.65e-03 | ||||
PDZ domain of connector enhancer of kinase suppressor of ras 1 (CNK1), CNK2, CNK3, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of CNK1 (also known as connector enhancer of KSR 1 (CNKSR1), CNK homolog protein 1, connector enhancer of KSR-like), CNK2 (also known as CNKSR2, CNK homolog protein 2), and CNK3 (also known as CNKSR3, CNK homolog protein 3, CNKSR family member 3, maguin-like). CNK proteins modulate Ras-mediated signaling, acting downstream of Ras as a scaffold for the Raf/MEK/ERK kinase cascade. They also modulate signaling mediated via Rho family small GTPases, through interactions with various guanine nucleotide exchange factors (GEFs) and GTPase activating proteins (GAPs), and modulate the insulin signaling pathway through interactions with the Arf guanine nucleotide exchange factors. CNK proteins also regulate cell proliferation and migration by acting as scaffolds for the PI3K/Akt and JNK signaling cascades. CNK2 plays a role in the molecular processes that govern morphology of the postsynaptic density (PSD), and influences subcellular localization of the regulatory NCK-interacting kinase TNIK. TNIK binds a region of CNK2 including the PDZ and the DUF domain; this region also binds the kinase MINK1. CNK2 may also influence the membrane localization of MINK1. CNK3 plays a part in transepithelial sodium transport; it coordinates assembly of an epithelial sodium channel (ENaC)-regulatory complex. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This CNK1-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged as beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta- strand F. Pssm-ID: 467230 [Multi-domain] Cd Length: 81 Bit Score: 38.36 E-value: 1.65e-03
|
||||||||
| Ubox | smart00504 | Modified RING finger domain; Modified RING finger domain, without the full complement of Zn2 ... |
15-77 | 1.67e-03 | ||||
Modified RING finger domain; Modified RING finger domain, without the full complement of Zn2+-binding ligands. Probable involvement in E2-dependent ubiquitination. Pssm-ID: 128780 [Multi-domain] Cd Length: 63 Bit Score: 37.98 E-value: 1.67e-03
|
||||||||
| RING-HC_PRT1-like | cd23132 | RING finger, HC subclass, found in Arabidopsis thaliana proteolysis 1 protein (PRT1) and ... |
15-57 | 1.69e-03 | ||||
RING finger, HC subclass, found in Arabidopsis thaliana proteolysis 1 protein (PRT1) and similar proteins; PRT1, also called RING-type E3 ubiquitin transferase PRT1, is an E3 ubiquitin-protein ligase that mediates ubiquitination and subsequent proteasomal degradation of target proteins. It functions in the N-end rule pathway of protein degradation, where it specifically recognizes and ubiquitinates proteins with an N-terminal bulky aromatic amino acid (Phe). It does not act on aliphatic hydrophobic and basic N-terminal residues (Arg or Leu) containing proteins. PRT1 contains a typical C3HC4-type RING-HC finger. Pssm-ID: 438494 [Multi-domain] Cd Length: 52 Bit Score: 37.40 E-value: 1.69e-03
|
||||||||
| PDZ_6 | pfam17820 | PDZ domain; This entry represents the PDZ domain from a wide variety of proteins. |
282-338 | 1.77e-03 | ||||
PDZ domain; This entry represents the PDZ domain from a wide variety of proteins. Pssm-ID: 436067 [Multi-domain] Cd Length: 54 Bit Score: 37.51 E-value: 1.77e-03
|
||||||||
| PDZ3_GRIP1-2-like | cd06684 | PDZ domain 3 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related ... |
446-490 | 1.83e-03 | ||||
PDZ domain 3 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR) binding proteins GRIP1 (ABP/GRIP2) and GRIP2, and related domains. GRIP1 and GRIP2 each have 7 PDZ domains. The interaction of GRIP1 and GRIP2 with GluA2/3 (AMPAR subunit) regulates AMPAR trafficking and synaptic targeting. GRIP1 has an essential role in regulating AMPAR trafficking during synaptic plasticity and learning and memory. GRIP1 and GRIP2 interact with a variety of other proteins associated with protein trafficking and internalization, for example GRIP1 also interacts with KIF5 (also known as kinesin 1), EphB receptors, scaffold protein liprin-alpha, and the rasGEF GRASP-1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This GRIP family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467172 [Multi-domain] Cd Length: 87 Bit Score: 38.39 E-value: 1.83e-03
|
||||||||
| PDZ_RapGEF2_RapGEF6-like | cd06755 | PDZ domain of Rap guanine nucleotide exchange factor 2 and Rap guanine nucleotide exchange ... |
446-482 | 1.85e-03 | ||||
PDZ domain of Rap guanine nucleotide exchange factor 2 and Rap guanine nucleotide exchange factor 6, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of Rap guanine nucleotide exchange factor 2 (RapGEF2, also named RA-GEF-1, PDZ-GEF1, CNrasGEF and nRapGEP) and Rap guanine nucleotide exchange factor 6 (RapGEF6, also named RA-GEF-2 and PDZ-GEF2). RapGEF2 and RapGEF6 constitute a subfamily of guanine nucleotide exchange factors (GEFs) for RAP small GTPases that is characterized by the possession of the PDZ and Ras/Rap-associating domains. They activate Rap small GTPases, by catalyzing the release of GDP from the inactive GDP-bound forms, thereby accelerating GTP loading to yield the active GTP-bound forms. The PDZ domain of RapGEF6 (also known as PDZ-GEF2) binds junctional adhesion molecule A (JAM-A). PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This RapGEF2 and RapGEF6 family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467237 [Multi-domain] Cd Length: 83 Bit Score: 38.40 E-value: 1.85e-03
|
||||||||
| zf-TRAF | pfam02176 | TRAF-type zinc finger; |
101-139 | 1.94e-03 | ||||
TRAF-type zinc finger; Pssm-ID: 280357 [Multi-domain] Cd Length: 60 Bit Score: 37.44 E-value: 1.94e-03
|
||||||||
| RING-H2_RNF139-like | cd16476 | RING finger, H2 subclass, found in RING finger proteins RNF139, RNF145, and similar proteins; ... |
18-53 | 2.07e-03 | ||||
RING finger, H2 subclass, found in RING finger proteins RNF139, RNF145, and similar proteins; RNF139, also known as translocation in renal carcinoma on chromosome 8 protein (TRC8), is an endoplasmic reticulum (ER)-resident multi-transmembrane protein that functions as a potent growth suppressor in mammalian cells, inducing G2/M arrest, decreased DNA synthesis and increased apoptosis. It is a tumor suppressor that has been implicated in a novel regulatory relationship linking the cholesterol/lipid biosynthetic pathway with cellular growth control. A mutation in RNF139 has been identified in families with hereditary renal (RCC) and thyroid cancers. RNF145 is an uncharacterized RING finger protein encoded by the RNF145 gene, which is expressed in T lymphocytes, and its expression is altered in acute myelomonocytic and acute promyelocytic leukemias. Although its biological function remains unclear, RNF145 shows high sequence similarity with RNF139. Both RNF139 and RNF145 contain a C3H2C3-type RING-H2 finger with possible E3-ubiquitin ligase activity. Pssm-ID: 438139 [Multi-domain] Cd Length: 41 Bit Score: 37.05 E-value: 2.07e-03
|
||||||||
| RING-HC_MID1 | cd16753 | RING finger, HC subclass, found in midline-1 (MID1) and similar proteins; MID1, also known as ... |
12-41 | 2.09e-03 | ||||
RING finger, HC subclass, found in midline-1 (MID1) and similar proteins; MID1, also known as midin, midline 1 RING finger protein, putative transcription factor XPRF, RING finger protein 59 (RNF59), or tripartite motif-containing protein 18 (TRIM18), is a microtubule-associated E3 ubiquitin-protein ligase implicated in epithelial-mesenchymal differentiation, cell migration and adhesion, and programmed cell death along specific regions of the ventral midline during embryogenesis. It monoubiquinates the alpha4 subunit of protein phosphatase 2A (PP2A), promoting proteosomal degradation of the catalytic subunit of PP2A (PP2Ac) and preventing the A and B subunits from forming an active complex. It promotes allergen and rhinovirus-induced asthma through the inhibition of PP2A activity. It is strongly upregulated in cytotoxic lymphocytes (CTLs) and directs lytic granule exocytosis and cytotoxicity of killer T cells. Loss-of-function mutations in MID1 lead to the human X-linked Opitz G/BBB (XLOS) syndrome characterized by defective midline development during embryogenesis. MID1 belongs to the C-I subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region, as well as a COS (carboxyl-terminal subgroup one signature) box, a fibronectin type III (FN3) domain, and a B30.2/SPRY (SplA and ryanodine receptor) domain positioned C-terminal to the RBCC domain. MID1 hetero-dimerizes in vitro with its paralog MID2. Pssm-ID: 438411 [Multi-domain] Cd Length: 72 Bit Score: 37.71 E-value: 2.09e-03
|
||||||||
| cpPDZ1_EcRseP-like | cd23082 | circularly permuted PDZ domain 1 (PDZ-N) of Escherichia coli Regulator of sigma-E protease ... |
448-479 | 2.18e-03 | ||||
circularly permuted PDZ domain 1 (PDZ-N) of Escherichia coli Regulator of sigma-E protease (RseP) and related domains; Permuted PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of ResP (also known as Site-2 protease RseP, and YaeL) and related domains. RseP is involved in the regulation of an extracytoplasmic stress response through the cleavage of membrane-spanning anti-stress-response transcription factor (anti-sigmaE) protein RseA; it cleaves the peptide bond between the critical alanine and cysteine in the transmembrane region of RseA, releasing the cytoplasmic domain of RseA with it associated sigmaE. RseP contains two tandem-arranged periplasmic PDZ domains (PDZ-N/PDZ1 and PDZ-C/PDZ2) which act to negatively regulate protease action on intact RseA; they serve as a size-exclusion filter which prevents the access of an intact RseA into the active site of RseP. PDZ domains usually bind in sequence-specific manner to short peptide sequences located at the C-terminal of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains and as well as those with circular permutations and domain swapping of beta-strands. The canonical PDZ domain contains six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. This RseP family PDZ domain is a circularly permuted PDZ domain which places both beta-strands A and B at the C-terminus. Another permutation exists in the PDZ superfamily which places beta-strand A at the C-terminus. Pssm-ID: 467639 [Multi-domain] Cd Length: 89 Bit Score: 38.12 E-value: 2.18e-03
|
||||||||
| PDZ1_GRIP1-2-like | cd06687 | PDZ domain 1 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related ... |
254-335 | 2.25e-03 | ||||
PDZ domain 1 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR) binding proteins GRIP1 (ABP/GRIP2) and GRIP2, and related domains. GRIP1 and GRIP2 each have 7 PDZ domains. The interaction of GRIP1 and GRIP2 with GluA2/3 (AMPAR subunit) regulates AMPAR trafficking and synaptic targeting. GRIP1 has an essential role in regulating AMPAR trafficking during synaptic plasticity and learning and memory. GRIP1 and GRIP2 interact with a variety of other proteins associated with protein trafficking and internalization, for example GRIP1 also interacts with KIF5 (also known as kinesin 1), EphB receptors, scaffold protein liprin-alpha, and the rasGEF GRASP-1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This GRIP family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467175 [Multi-domain] Cd Length: 83 Bit Score: 38.16 E-value: 2.25e-03
|
||||||||
| PDZ_RGS3-like | cd06711 | PDZ domain of regulator of G-protein signaling 3 (RGS3), and related domains; PDZ (PSD-95 ... |
254-337 | 2.69e-03 | ||||
PDZ domain of regulator of G-protein signaling 3 (RGS3), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of RGS3, and related domains. RGS3 down-regulates GPCR signal transduction by increasing the GTPase activity of G-protein alpha subunits, thereby driving G-proteins into their inactive GDP-bound form. It downregulates G-protein-mediated release of inositol phosphates and activation of MAP kinases. In Eph/ephrin signaling, RGS3 binds via its PDZ domain to the cytoplasmic C terminus of Eph receptor tyrosine kinase EphB. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This RGS3-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467195 [Multi-domain] Cd Length: 77 Bit Score: 37.75 E-value: 2.69e-03
|
||||||||
| RING-H2_RNF165 | cd16682 | RING finger, H2 subclass, found in RING finger protein 165 (RNF165) and similar proteins; ... |
9-57 | 2.86e-03 | ||||
RING finger, H2 subclass, found in RING finger protein 165 (RNF165) and similar proteins; RNF165, also known as Arkadia-like 2, Arkadia2, or Ark2C, is an E3 ubiquitin ligase with homology to the C-terminal half of RNF111. It is expressed specifically in the nervous system, and can serve to amplify neuronal responses to specific signals. It acts as a positive regulator of bone morphogenetic protein (BMP)-Smad signaling that is involved in motor neuron (MN) axon elongation. RNF165 contains two serine rich domains, a nuclear localization signal, an NRG-TIER domain, and a C-terminal C3H2C3-type RING-H2 finger that is responsible for the enhancement of BMP-Smad1/5/8 signaling in the spinal cord. Pssm-ID: 438344 [Multi-domain] Cd Length: 59 Bit Score: 36.98 E-value: 2.86e-03
|
||||||||
| cpPDZ_BsHtra-like | cd06781 | circularly permuted PDZ domain of Bacillus subtilis HtrA-type serine proteases HtrA, HtrB, and ... |
441-483 | 2.90e-03 | ||||
circularly permuted PDZ domain of Bacillus subtilis HtrA-type serine proteases HtrA, HtrB, and YyxA and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of Bacillus subtilis HtrA/YkdA, HtrB/YvtA and YyxA/YycK, and related domains. HtrA-type serine proteases participate in folding and degradation of aberrant proteins, and in processing and maturation of native proteins. HtrA, HtrB, and YyxA have a single transmembrane domain at the N-terminus and a PDZ domain at the C-terminus. Expression of htrA and htrB genes is induced both by heat shock and by secretion stress (by a common) mechanism; yyxA is neither heat shock nor secretion stress inducible. HtrA and HtrB may have overlapping cellular functions; YyxA may have a cellular function distinct from the other two proteases or have the same function but under different conditions. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains and as well as those with circular permutations and domain swapping of beta-strands. The canonical PDZ domain contains six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. This BsHtrA-like PDZ domain is a circularly permuted PDZ domain which places beta-strand A on the C-terminus. Another permutation exists in the PDZ superfamily which places both beta-strands A and B on the C-terminus. Pssm-ID: 467622 [Multi-domain] Cd Length: 98 Bit Score: 38.00 E-value: 2.90e-03
|
||||||||
| PDZ2_APBA1_3-like | cd06793 | PDZ domain 2 of amyloid-beta A4 precursor protein-binding family A member 1 (APBA1), APBA2, ... |
267-327 | 2.99e-03 | ||||
PDZ domain 2 of amyloid-beta A4 precursor protein-binding family A member 1 (APBA1), APBA2, APBA3, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 2 of APBA1, APBA2, APBA3, and related domains. The APBA/X11/Mint protein family includes three members: neuron specific APBA1 (also known as X11alpha and Mint1) and APBA2 (also known as X11beta and Mint2), and the ubiquitously expressed APBA3 (also known as X12gamma and Mint3). They are involved in regulating neuronal signaling, trafficking, and plasticity. They contain two PDZ domains (PDZ1 and PDZ2) which bind a variety of proteins: Arf GTPases (APBA1 and APBA2 PDZ2) and neurexin (APBA1 and APBA2 PDZ1 and 2) which are involved in vesicle docking and exocytosis; alpha1B subunit of N-type Ca2+ channel (APBA1 PDZ1) that is involved in ion channels; KIF17 (APBA1 PDZ1) that is involved in transport and traffic; and Alzheimer's disease related proteins, APP (APBA3 PDZ2), CCS (APBA1 PDZ2), NF-kappa-B/p65 (APBA2 PDZ2), presenilin-1 (APBA1 and APBA2 PDZ1 and PDZ2). PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This APBA1,3-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged as beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467255 [Multi-domain] Cd Length: 78 Bit Score: 37.38 E-value: 2.99e-03
|
||||||||
| RING-HC_TRAF1-like | cd23125 | RING finger, HC subclass, found in tumor necrosis factor (TNF) receptor-associated factor 1 ... |
18-58 | 3.07e-03 | ||||
RING finger, HC subclass, found in tumor necrosis factor (TNF) receptor-associated factor 1 (TRAF1)-like and similar proteins; TRAF1, also known as Epstein-Barr virus-induced protein 6 (EBI6), is an adapter molecule that regulates the activation of NF-kappa-B and JNK. It plays a role in the regulation of cell survival and apoptosis. The heterotrimer formed by TRAF1 and TRAF2 is part of an E3 ubiquitin-protein ligase complex that promotes ubiquitination of target proteins, such as MAP3K14. The TRAF1/TRAF2 complex recruits the antiapoptotic E3 protein-ubiquitin ligases BIRC2 and BIRC3 to TNFRSF1B/TNFR2. This subfamily corresponds a group of TRAF1-like proteins that contains an N-terminal domain with a typical C3HC4-type RING-HC finger. Pssm-ID: 438487 [Multi-domain] Cd Length: 51 Bit Score: 36.73 E-value: 3.07e-03
|
||||||||
| RING-HC_RING1-like | cd16531 | RING finger, HC subclass, found in really interesting new gene proteins RING1, RING2 and ... |
15-63 | 3.14e-03 | ||||
RING finger, HC subclass, found in really interesting new gene proteins RING1, RING2 and similar proteins; RING1, also known as polycomb complex protein RING1, RING finger protein 1 (RNF1), or RING finger protein 1A (RING1A), is a transcriptional repressor that is associated with the Polycomb group (PcG) protein complex involved in stable repression of gene activity. RING2, also known as huntingtin-interacting protein 2-interacting protein 3, HIP2-interacting protein 3, protein DinG, RING finger protein 1B (RING1B), RING finger protein 2 (RNF2), or RING finger protein BAP-1, is an E3 ubiquitin-protein ligase that interacts with both nucleosomal DNA and an acidic patch on histone H4 to achieve the specific monoubiquitination of K119 on histone H2A (H2AK119ub), thereby playing a central role in histone code and gene regulation. Both RING1 and RING2 are core components of polycomb repressive complex 1 (PRC1) that functions as an E3-ubuiquitin ligase transferring the mono-ubuiquitin mark to the C-terminal tail of Histone H2A at K118/K119. PRC1 is also capable of chromatin compaction, a function not requiring histone tails, and this activity appears important in gene silencing. RING2 acts as the main E3 ubiquitin ligase on histone H2A of the PRC1 complex, while RING1 may rather act as a modulator of RNF2/RING2 activity. Members of this family contain a C3HC4-type RING-HC finger. Pssm-ID: 438193 [Multi-domain] Cd Length: 66 Bit Score: 37.25 E-value: 3.14e-03
|
||||||||
| RING-HC_TRIM8_C-V | cd16580 | RING finger, HC subclass, found in tripartite motif-containing protein 8 (TRIM8) and similar ... |
15-72 | 3.54e-03 | ||||
RING finger, HC subclass, found in tripartite motif-containing protein 8 (TRIM8) and similar proteins; TRIM8, also known as glioblastoma-expressed RING finger protein (GERP) or RING finger protein 27 (RNF27), is a probable E3 ubiquitin-protein ligase that may promote proteasomal degradation of suppressor of cytokine signaling 1 (SOCS1) and further regulate interferon-gamma signaling. It functions as a new p53 modulator that stabilizes p53 impairing its association with MDM2 and inducing the reduction of cell proliferation. TRIM8 deficit dramatically impairs p53 stabilization and activation in response to chemotherapeutic drugs. TRIM8 also modulates tumor necrosis factor-alpha (TNFalpha) and interleukin-1beta (IL-1beta)-triggered nuclear factor-kappaB (NF- kappa B) activation by targeting transforming growth factor beta (TGFbeta) activated kinase 1 (TAK1) for K63-linked polyubiquitination. Moreover, TRIM8 modulates translocation of phosphorylated STAT3 into the nucleus through interaction with Hsp90beta and consequently regulates transcription of Nanog in embryonic stem cells. It also interacts with protein inhibitor of activated STAT3 (PIAS3), which inhibits IL-6-dependent activation of STAT3. TRIM8 belongs to the C-V subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil domain, as well as an uncharacterized region positioned C-terminal to the RBCC domain. The coiled coil domain is required for homodimerization and the region immediately C-terminal to the RING motif is sufficient to mediate the interaction with SOCS1. Pssm-ID: 438242 [Multi-domain] Cd Length: 67 Bit Score: 37.18 E-value: 3.54e-03
|
||||||||
| zf-RING_5 | pfam14634 | zinc-RING finger domain; |
17-57 | 3.55e-03 | ||||
zinc-RING finger domain; Pssm-ID: 434085 [Multi-domain] Cd Length: 43 Bit Score: 36.25 E-value: 3.55e-03
|
||||||||
| cpPDZ_BsHtra-like | cd06781 | circularly permuted PDZ domain of Bacillus subtilis HtrA-type serine proteases HtrA, HtrB, and ... |
279-337 | 3.55e-03 | ||||
circularly permuted PDZ domain of Bacillus subtilis HtrA-type serine proteases HtrA, HtrB, and YyxA and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of Bacillus subtilis HtrA/YkdA, HtrB/YvtA and YyxA/YycK, and related domains. HtrA-type serine proteases participate in folding and degradation of aberrant proteins, and in processing and maturation of native proteins. HtrA, HtrB, and YyxA have a single transmembrane domain at the N-terminus and a PDZ domain at the C-terminus. Expression of htrA and htrB genes is induced both by heat shock and by secretion stress (by a common) mechanism; yyxA is neither heat shock nor secretion stress inducible. HtrA and HtrB may have overlapping cellular functions; YyxA may have a cellular function distinct from the other two proteases or have the same function but under different conditions. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains and as well as those with circular permutations and domain swapping of beta-strands. The canonical PDZ domain contains six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. This BsHtrA-like PDZ domain is a circularly permuted PDZ domain which places beta-strand A on the C-terminus. Another permutation exists in the PDZ superfamily which places both beta-strands A and B on the C-terminus. Pssm-ID: 467622 [Multi-domain] Cd Length: 98 Bit Score: 38.00 E-value: 3.55e-03
|
||||||||
| RING-HC_MID2 | cd16754 | RING finger, HC subclass, found in midline-2 (MID2) and similar proteins; MID2, also known as ... |
12-41 | 3.58e-03 | ||||
RING finger, HC subclass, found in midline-2 (MID2) and similar proteins; MID2, also known as midin-2, midline defect 2, RING finger protein 60 (RNF60), or tripartite motif-containing protein 1 (TRIM1), is a probable E3 ubiquitin-protein ligase and is highly related to MID1 that associates with cytoplasmic microtubules along their length and throughout the cell cycle. Like MID1, MID2 associates with the microtubule network and may at least partially compensate for the loss of MID1. Both MID1 and MID2 interacts with Alpha 4, which is a regulatory subunit of PP2-type phosphatases, such as PP2A, and an integral component of the rapamycin-sensitive signaling pathway. MID2 can also substitute for MID1 to control exocytosis of lytic granules in cytotoxic T cells. Loss-of-function mutations in MID2 lead to the human X-linked intellectual disability (XLID). MID2 belongs to the C-I subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region, as well as a COS (carboxy-terminal subgroup one signature) box, a fibronectin type III (FN3) domain, and a B30.2/SPRY (SplA and ryanodine receptor) domain positioned C-terminal to the RBCC domain. MID2 hetero-dimerizes in vitro with its paralog MID1. Pssm-ID: 438412 [Multi-domain] Cd Length: 70 Bit Score: 37.27 E-value: 3.58e-03
|
||||||||
| PDZ4_MAGI-1_3-like | cd06734 | PDZ domain 4 of membrane-associated guanylate kinase inverted 1 (MAGI-1), MAGI-2, and MAGI-3, ... |
445-480 | 3.68e-03 | ||||
PDZ domain 4 of membrane-associated guanylate kinase inverted 1 (MAGI-1), MAGI-2, and MAGI-3, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 4 of MAGI1, 2, 3 (MAGI is also known as Membrane-associated guanylate kinase, WW and PDZ domain-containing protein) and related domains. MAGI proteins have been implicated in the control of cell migration and invasion through altering the activity of phosphatase and tensin homolog (PTEN) and modulating Akt signaling. Four MAGI proteins have been identified (MAGI1-3 and MAGIX). MAGI1-3 have 6 PDZ domains and bind to the C-terminus of PTEN via their PDZ2 domain. MAGIX has a single PDZ domain that is related to MAGI1-3 PDZ domain 5. Other binding partners for MAGI1 include JAM4, C-terminal tail of high risk HPV-18 E6, megalin, TRAF6, Kir4.1 (basolateral K+ channel subunit), and cadherin 23; for MAGI2, include DASM1, dendrin, axin, beta- and delta-catenin, neuroligin, hyperpolarization-activated cation channels, beta1-adrenergic receptors, NMDA receptor, and TARPs; and for MAGI3 includes LPA2. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MAGI family PDZ4 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as beta-strands A, -B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467216 [Multi-domain] Cd Length: 84 Bit Score: 37.59 E-value: 3.68e-03
|
||||||||
| COG3975 | COG3975 | Predicted metalloprotease, contains C-terminal PDZ domain [General function prediction only]; |
256-338 | 4.17e-03 | ||||
Predicted metalloprotease, contains C-terminal PDZ domain [General function prediction only]; Pssm-ID: 443174 [Multi-domain] Cd Length: 591 Bit Score: 40.96 E-value: 4.17e-03
|
||||||||
| RING-HC_PCGF | cd16525 | RING finger, HC subclass, found in Polycomb Group RING finger homologs (PCGF1, 2, 3, 4, 5 and ... |
16-57 | 4.19e-03 | ||||
RING finger, HC subclass, found in Polycomb Group RING finger homologs (PCGF1, 2, 3, 4, 5 and 6), and similar proteins; This subfamily includes six Polycomb Group (PcG) RING finger homologs (PCGF1/NSPc1, PCGF2/Mel-18, PCGF3, PCGF4/BMI1, PCGF5, and PCGF6/MBLR) that use epigenetic mechanisms to maintain or repress expression of their target genes. They were first discovered in fruit flies and are well known for silencing Hox genes through modulation of chromatin structure during embryonic development. PCGF homologs play important roles in cell proliferation, differentiation, and tumorigenesis. They all have been found to associate with ring finger protein 2 (RNF2). The RNF2-PCGF heterodimer is catalytically competent as an E3 ubiquitin transferase and is the scaffold for the assembly of additional components. Moreover, PCGF homologs are critical components in the assembly of distinct Polycomb Repression Complex 1 (PRC1) related complexes which is involved in the maintenance of gene repression and which target different genes through distinct mechanisms. The Drosophila PRC1 core complex is formed by the Polycomb (Pc), Polyhomeotic (Ph), Posterior sex combs (Psc), and Sex combs extra (Sce, also known as Ring) subunits. In mammals, the composition of PRC1 is much more diverse and varies depending on the cellular context. All PRC1 complexes contain homologs of the Drosophila Ring protein. Ring1A/RNF1 and Ring1B/RNF2 are E3 ubiquitin ligases that mark lysine 119 of histone H2A with a single ubiquitin group (H2AK119ub). Mammalian homologs of the Drosophila Psc protein, such as PCGF2/Mel-18 or PCGF4/BMI1, regulate PRC1 enzymatic activity. PRC1 complexes can be divided into at least two classes according to the presence or absence of CBX proteins, which are homologs of Drosophila Pc. Canonical PRC1 complexes contain CBX proteins that recognize and bind H3K27me3, the mark deposited by PRC2. Therefore, canonical PRC1 complexes and PRC2 can act together to repress gene transcription and maintain this repression through cell division. Non-canonical PRC1 complexes, containing RYBP (together with additional proteins, such as L3mbtl2 or Kdm2b) rather than the CBX proteins have recently been described in mammals. PCGF homologs contain a C3HC4-type RING-HC finger. Pssm-ID: 438188 [Multi-domain] Cd Length: 42 Bit Score: 36.05 E-value: 4.19e-03
|
||||||||
| Prok-RING_4 | pfam14447 | Prokaryotic RING finger family 4; RING finger family domain found sporadically in bacteria. ... |
18-62 | 4.38e-03 | ||||
Prokaryotic RING finger family 4; RING finger family domain found sporadically in bacteria. The finger is fused to an N-terminal alpha-helical domain, ROT/Trove-like repeats and a C-terminal TerD domain. The architecture suggests a possible role in an RNA-processing complex. Pssm-ID: 433959 [Multi-domain] Cd Length: 46 Bit Score: 36.25 E-value: 4.38e-03
|
||||||||
| PDZ1_PDZ3_bazooka-like | cd06665 | PDZ domain 1 and PDZ domain 3 of Drosophila bazooka (DmPar3), and related domains; PDZ (PSD-95 ... |
431-492 | 4.69e-03 | ||||
PDZ domain 1 and PDZ domain 3 of Drosophila bazooka (DmPar3), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 and 3 of Drosophila bazooka (DmPar3), and related domains. The Par complex comprises atypical protein kinase C (aPKC) and two scaffolding proteins, Par3 (Bazooka (Baz) in Drosophila) and Par6; bazooka (DmPar3) has three central PDZ domains. Both PDZ1 and PDZ3 domains, but not PDZ2, in bazooka (DmPar3) engage in a canonical interaction with the PDZ domain-binding motif in Par6. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This bazooka-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467154 [Multi-domain] Cd Length: 87 Bit Score: 37.34 E-value: 4.69e-03
|
||||||||
| PDZ1_ZO1-like | cd06727 | PDZ domain 1 of Zonula Occludens-1 (ZO-1), homologs ZO-2 and ZO-3, and related domains; PDZ ... |
444-486 | 4.74e-03 | ||||
PDZ domain 1 of Zonula Occludens-1 (ZO-1), homologs ZO-2 and ZO-3, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of ZO-1, -2, -3 and related domains. Zonula occludens proteins (ZO-1, ZO-2, ZO-3) are multi-PDZ domain proteins involved in the maintenance and biogenesis of multi-protein networks at the cytoplasmic surface of intercellular contacts in epithelial and endothelial cells. They have three N-terminal PDZ domains, PDZ1-3, followed by a Src homology-3 (SH3) domain and a guanylate kinase (GuK)-like domain. Among protein-protein interactions for all ZO proteins is the binding of the first PDZ domain (PDZ1) to the C-termini of claudins, and the homo- and hetero-dimerization of ZO-proteins via their second PDZ domain (PDZ2), which takes place by symmetrical domain swapping of the first two beta-strands of PDZ2. At the cell level, ZO-1 and ZO-2 are involved in polarity maintenance, gene transcription, cell proliferation, and tumor cell metastasis. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This ZO family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467209 [Multi-domain] Cd Length: 87 Bit Score: 37.25 E-value: 4.74e-03
|
||||||||
| RING-Ubox_LubX-like_rpt1 | cd23149 | first U-box domain, a modified RING finger, found in Legionella pneumophila U-box protein LubX ... |
16-65 | 4.77e-03 | ||||
first U-box domain, a modified RING finger, found in Legionella pneumophila U-box protein LubX and similar proteins; LubX, also called RING-type E3 ubiquitin transferase LubX, is part of the large arsenal of effectors in Legionella pneumophila that are translocated into the host cytosol during infection. LubX acts as an E3 ubiquitin-protein ligase (EC 2.3.2.27) that interferes with the host's ubiquitination pathway. LubX contains two RING-like U-box domains. U-box 1 is critical to the ubiquitin ligase activity, and U-box 2 mediates interaction with host target. This model corresponds to the first one. Pssm-ID: 438511 [Multi-domain] Cd Length: 55 Bit Score: 36.31 E-value: 4.77e-03
|
||||||||
| PDZ_SIPA1-like | cd06745 | PDZ domain of signal-induced proliferation-associated protein 1 (SIPA1), and related domains; ... |
250-336 | 4.80e-03 | ||||
PDZ domain of signal-induced proliferation-associated protein 1 (SIPA1), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of SIPA1, and related domains. The Rap-GTPase activating protein SIPA1 (also known as GTPase-activating protein Spa-1, p130 SPA1) is a metastasis promoter; a polymorphism in a region of the Sipa1 gene encoding the PDZ domain is associated with metastasis. The SIPA1 PDZ domain binds ribosomal RNA processing 1 homolog B (Rrp1b). SIPA1 also forms a complex with water channel aquaporin-2 (AQP2) and plays a role in trafficking of AQP2, targeted positioning of which strictly regulates body water homeostasis; the SIPA1 PDZ domain binds AQP2. Rrp1b or AQP2 binding inhibits the RapGAP activity of SIPA1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This SIPA1-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged as beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta- strand F. Pssm-ID: 467227 [Multi-domain] Cd Length: 73 Bit Score: 36.87 E-value: 4.80e-03
|
||||||||
| PDZ_shroom2_3_4-like | cd06750 | PDZ domain of shroom2, shroom3, shroom4, and related domains; PDZ (PSD-95 (Postsynaptic ... |
260-338 | 4.81e-03 | ||||
PDZ domain of shroom2, shroom3, shroom4, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of shroom2, shroom3, shroom4, and related domains. Shroom family proteins shroom2 (also known as apical-like protein; protein APXL), shroom3 (also known as shroom-related protein), and shroom4 (also known as second homolog of apical protein) are essential regulators of cell morphology during animal development; they regulate cell architecture by directing the subcellular distribution and activation of Rho kinase (ROCK), which results in the localized activation of non-muscle myosin. The interaction between shroom and ROCK is mediated by the shroom domain 2 (SD2). PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This shroom2,3,4-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged as beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467232 [Multi-domain] Cd Length: 82 Bit Score: 36.93 E-value: 4.81e-03
|
||||||||
| zf-C3HC4_3 | pfam13920 | Zinc finger, C3HC4 type (RING finger); |
15-57 | 5.11e-03 | ||||
Zinc finger, C3HC4 type (RING finger); Pssm-ID: 464042 [Multi-domain] Cd Length: 50 Bit Score: 36.20 E-value: 5.11e-03
|
||||||||
| RING-HC_TRIM13_C-V | cd16762 | RING finger, HC subclass, found in tripartite motif-containing protein 13 (TRIM13) and similar ... |
15-42 | 5.26e-03 | ||||
RING finger, HC subclass, found in tripartite motif-containing protein 13 (TRIM13) and similar proteins; TRIM13, also known as B-cell chronic lymphocytic leukemia tumor suppressor Leu5, leukemia-associated protein 5, putative tumor suppressor RFP2, RING finger protein 77 (RNF77), or Ret finger protein 2, is an endoplasmic reticulum (ER) membrane anchored E3 ubiquitin-protein ligase that interacts with proteins localized to the ER, including valosin-containing protein (VCP), a protein indispensable for ER-associated degradation (ERAD). It also targets the known ER proteolytic substrate CD3-delta, but not the N-end rule substrate Ub-R-YFP (yellow fluorescent protein) for degradation. Moreover, TRIM13 regulates ubiquitination and degradation of NEMO to suppress tumor necrosis factor (TNF) induced nuclear factor-kappaB (NF- kappa B) activation. It is also involved in NF-kappaB p65 activation and nuclear factor of activated T-cells (NFAT)-dependent activation of c-Rel upon T-cell receptor engagement. Furthermore, TRIM13 negatively regulates melanoma differentiation-associated gene 5 (MDA5)-mediated type I interferon production. It also regulates caspase-8 ubiquitination, translocation to autophagosomes, and activation during ER stress induced cell death. Meanwhile, TRIM13 enhances ionizing radiation-induced apoptosis by increasing p53 stability and decreasing AKT kinase activity through MDM2 and AKT degradation. TRIM13 belongs to the C-V subclass of the TRIM (tripartite motif) family of proteins that are defined by an N-terminal RBCC (RING, Bbox, and coiled coil) domain, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region. In addition, TRIM13 contains a C-terminal transmembrane domain. Pssm-ID: 438418 [Multi-domain] Cd Length: 56 Bit Score: 36.05 E-value: 5.26e-03
|
||||||||
| RING-HC_RNF39 | cd16592 | RING finger, HC subclass, found in RING finger protein 39 (RNF39) and similar proteins; RNF39, ... |
18-42 | 5.38e-03 | ||||
RING finger, HC subclass, found in RING finger protein 39 (RNF39) and similar proteins; RNF39, also called protein HZFw, may play a role in prolonged long term-potentiation (LTP) maintenance. It is involved in the etiology of Behcet's disease (BD). It may also be involved in HIV-1 replication. RNF39 acts as an E3 ubiquitin ligase that inhibits retinoic acid-inducible gene-I (RIG-I)-like receptor (RLR) pathways by mediating K48-linked ubiquitination and proteasomal degradation of DDX3X (DEAD-box RNA helicase 3, X-linked). RNF39 contains a typical C3HC4-type RING-HC finger. Pssm-ID: 438254 [Multi-domain] Cd Length: 58 Bit Score: 36.27 E-value: 5.38e-03
|
||||||||
| RING-HC_RAD18 | cd16529 | RING finger, HC subclass, found in postreplication repair protein RAD18 and similar proteins; ... |
12-65 | 5.39e-03 | ||||
RING finger, HC subclass, found in postreplication repair protein RAD18 and similar proteins; RAD18, also known as HR18 or RING finger protein 73 (RNF73), is an E3 ubiquitin-protein ligase involved in post replication repair of UV-damaged DNA via its recruitment to stalled replication forks. It associates to the E2 ubiquitin conjugating enzyme UBE2B to form the UBE2B-RAD18 ubiquitin ligase complex involved in mono-ubiquitination of DNA-associated PCNA on K164. It also interacts with another E2 ubiquitin conjugating enzyme RAD6 to form a complex that monoubiquitinates proliferating cell nuclear antigen at stalled replication forks in DNA translesion synthesis. Moreover, Rad18 is a key factor in double-strand break DNA damage response (DDR) pathways via its association with K63-linked polyubiquitylated chromatin proteins. It can function as a mediator for DNA damage response signals to activate the G2/M checkpoint in order to maintain genome integrity and cell survival after ionizing radiation (IR) exposure. RAD18 contains a C3HC4-type RING-HC finger, a ubiquitin-binding zinc finger domain (UBZ), a SAP (SAF-A/B, Acinus and PIAS) domain, and a RAD6-binding domain (R6BD). Pssm-ID: 438192 [Multi-domain] Cd Length: 54 Bit Score: 36.13 E-value: 5.39e-03
|
||||||||
| PDZ4_MUPP1-like | cd06668 | PDZ domain 4 of multi-PDZ-domain protein 1 (MUPP1) and PATJ (protein-associated tight junction) ... |
282-325 | 5.42e-03 | ||||
PDZ domain 4 of multi-PDZ-domain protein 1 (MUPP1) and PATJ (protein-associated tight junction) and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 4 of MUPP1 and PATJ, and related domains. MUPP1 and PATJ serve as scaffolding proteins linking different proteins and protein complexes involved in the organization of tight junctions and epithelial polarity. MUPP1 contains an L27 (Lin-2 and Lin-7 binding) domain and 13 PDZ domains. PATJ (also known as INAD-like) contains an L27 domain and ten PDZ domains. MUPP1 and PATJ share several binding partners, including junctional adhesion molecules (JAM), zonula occludens (ZO)-3, Pals1 (protein associated with Lin-7), Par (partitioning defective)-6 proteins, and nectins (adherence junction adhesion molecules). PATJ lacks 3 PDZ domains seen in MUPP1: PDZ6, 9, and 13; consequently, MUPP1 PDZ7 and 8 align with PATJ PDZ6 and 7; and MUPP1 PDZ domains 10-12 align with PATJ PDZ domains 8-10. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MUPP1-like family PDZ4 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F Pssm-ID: 467156 [Multi-domain] Cd Length: 88 Bit Score: 37.28 E-value: 5.42e-03
|
||||||||
| RING-HC_TRAF3 | cd16640 | RING finger, HC subclass, found in tumor necrosis factor (TNF) receptor-associated factor 3 ... |
17-42 | 5.44e-03 | ||||
RING finger, HC subclass, found in tumor necrosis factor (TNF) receptor-associated factor 3 (TRAF3) and similar proteins; TRAF3, also known as CAP-1, CD40 receptor-associated factor 1 (CRAF1), CD40-binding protein (CD40BP), or LMP1-associated protein 1 (LAP1), is a member of the TRAF protein family, which mainly functions in the immune system, where it mediates signaling through tumor necrosis factor receptors (TNFRs) and interleukin-1/Toll-like receptors (IL-1/TLRs). It also plays a unique cell type-specific and critical role in the restraint of B-cell homeostatic survival, a role with important implications for both B-cell differentiation and the pathogenesis of B-cell malignancies. TRAF3 differentially regulates differentiation of specific T cell subsets. It is required for iNKT cell development, restrains Treg cell development in the thymus, and plays an essential role in the homeostasis of central memory CD8+ T cells. TRAF3 contains an N-terminal domain with a typical C3HC4-type RING-HC finger and several zinc fingers, and a C-terminal TRAF domain that comprises a coiled coil domain, and a conserved TRAF-C domain. Pssm-ID: 438302 [Multi-domain] Cd Length: 42 Bit Score: 35.64 E-value: 5.44e-03
|
||||||||
| mRING-H2-C3H3C2_Mio-like | cd16488 | Modified RING finger, H2 subclass (C3H3C2-type), found in WD repeat-containing protein mio and ... |
17-56 | 5.55e-03 | ||||
Modified RING finger, H2 subclass (C3H3C2-type), found in WD repeat-containing protein mio and its homologs; This subfamily contains Mio, WDR24, WDR59, and their counterparts Sea4, Sea2, and Sea3 from yeast, respectively. Mio/Sea4, Sea2/WDR24, and Sea3/WDR59 are components of the GATOR2 complex, which also includes another two subunits, Seh1and Sec13. GATOR2 and GATOR1, which is composed of three subunits, DEPDC5, Nprl2, and Nprl3, form the Rag-interacting complex GATOR (GAP Activity Towards Rags). Inhibition of GATOR1 subunits makes mTORC1 signaling resistant to amino acid deprivation. In contrast, inhibition of GATOR2 subunits suppresses mTORC1 signaling and GATOR2 negatively regulates DEPDC5. All subfamily members contain an N-terminal WD40 domain and a C-terminal RING-H2 finger with an unusual arrangement of zinc-coordinating residues. The cysteines and histidines in the RING-H2 finger are arranged as a modified C3H3C2-type, rather than the canonical C3H2C3-type. Pssm-ID: 438151 [Multi-domain] Cd Length: 44 Bit Score: 35.77 E-value: 5.55e-03
|
||||||||
| RING-HC_TRAF2 | cd16639 | RING finger, HC subclass, found in tumor necrosis factor (TNF) receptor-associated factor 2 ... |
18-56 | 5.64e-03 | ||||
RING finger, HC subclass, found in tumor necrosis factor (TNF) receptor-associated factor 2 (TRAF2) and similar proteins; TRAF2, also known as tumor necrosis factor type 2 receptor-associated protein 3, is an E3 ubiquitin-protein ligase that was identified as a 75 kDa tumor necrosis factor receptor (TNF-R2)-associated signaling protein. It interacts with members of the TNF receptor superfamily and connects the receptors to downstream signaling proteins. It also mediates K63-linked polyubiquitination of RIP1, a kinase pivotal in TNFalpha-induced NF-kappaB activation. Moreover, TRAF2 regulates peripheral CD8(+) T-cell and NKT-cell homeostasis by modulating sensitivity to IL-15. It also acts as an important biological suppressor of necroptosis. It inhibits TNF-related apoptosis inducing ligand (TRAIL)- and CD95L-induced apoptosis and necroptosis. TRAF2 contains an N-terminal domain with a typical C3HC4-type RING-HC finger and several zinc fingers, and a C-terminal TRAF domain that comprises a coiled coil domain and a conserved TRAF-C domain. Pssm-ID: 438301 [Multi-domain] Cd Length: 43 Bit Score: 35.90 E-value: 5.64e-03
|
||||||||
| RING-H2_synoviolin | cd16479 | RING finger, H2 subclass, found in synoviolin and similar proteins; Synoviolin, also known as ... |
32-57 | 5.87e-03 | ||||
RING finger, H2 subclass, found in synoviolin and similar proteins; Synoviolin, also known as synovial apoptosis inhibitor 1 (Syvn1), Hrd1, or Der3, is an endoplasmic reticulum (ER)-anchoring E3 ubiquitin ligase that functions as a suppressor of ER stress-induced apoptosis and plays a role in homeostasis maintenance. It also targets tumor suppressor gene p53 for proteasomal degradation, suggesting crosstalk between ER associated degradation (ERAD) and p53 mediated apoptotic pathway under ER stress. Moreover, synoviolin controls body weight and mitochondrial biogenesis through negative regulation of the thermogenic coactivator peroxisome proliferator-activated receptor coactivator (PGC)-1beta. It upregulates amyloid beta production by targeting a negative regulator of gamma-secretase, Retention in endoplasmic reticulum 1 (Rer1), for degradation. It is also involved in the degradation of endogenous immature nicastrin, and affects amyloid beta-protein generation. Moreover, synoviolin is highly expressed in rheumatoid synovial cells and may be involved in the pathogenesis of rheumatoid arthritis (RA). It functions as an anti-apoptotic factor that is responsible for the outgrowth of synovial cells during the development of RA. It promotes inositol-requiring enzyme 1 (IRE1) ubiquitination and degradation in synovial fibroblasts with collagen-induced arthritis. Furthermore, the upregulation of synoviolin may represent a protective response against neurodegeneration in Parkinson's disease (PD). In addition, synoviolin is involved in liver fibrogenesis. Synoviolin contains a C3H2C2-type RING-H2 finger. Pssm-ID: 438142 [Multi-domain] Cd Length: 43 Bit Score: 35.79 E-value: 5.87e-03
|
||||||||
| PDZ_MPP1-like | cd10830 | PDZ domain of membrane palmitoylated protein1 (MPP1), and related domains; PDZ (PSD-95 ... |
283-336 | 5.97e-03 | ||||
PDZ domain of membrane palmitoylated protein1 (MPP1), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of MPP1, and related domains. MPP1 (also known as MAGUK p55 subfamily member 1, erythrocyte membrane protein p55, EMP55) is a membrane-associated guanylate kinase (MAGUK)-like protein which forms a complex with protein 4.1 and glycophorin C (GPC) at the cytoplasmic face of the plasma membrane; this complex is essential for cytoskeleton-membrane linkage in erythrocytes and many non-erythroid cells, and participates in the determination of membrane stability and cell shape. MPP1, by interacting with various scaffold proteins and cytoskeletal proteins in the postsynaptic density, also plays an important role in organizing synaptic and non-synaptic structures. MPP1 is also a component of the Crumbs protein complex in the mammalian retina and may link the Usher protein network and the Crumbs protein complex. The MPP1 PDZ domain binding partners include GPC, ABCC4, and CADM1/Necl-2/SynCAM1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MPP1-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467266 [Multi-domain] Cd Length: 81 Bit Score: 36.77 E-value: 5.97e-03
|
||||||||
| PDZ3_FL-whirlin-like | cd06742 | PDZ domain 3 of the full-length isoform of whirlin, PDZ domain 1 of the short isoform of ... |
283-326 | 6.21e-03 | ||||
PDZ domain 3 of the full-length isoform of whirlin, PDZ domain 1 of the short isoform of whirlin, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 3 of the full-length isoform of whirlin, PDZ domain 1 of the short isoform of whirlin, and related domains. Whirlin is an essential protein for developmental pathways in photoreceptor cells of the retina and hair cells of the inner ear. The full-length whirlin isoform has two harmonin N-like domains, three PDZ domains, a proline-rich region, and a PDZ-binding motif. Whirlin isoforms may form different complexes at the periciliary membrane complex (PMC) in photoreceptors, and the stereociliary tip and base in inner ear hair cells. It interacts with ADGRV1 and usherin at the PMC; with SANS and RpgrORF15 at the connecting cilium in photoreceptors; with EPS8, MYO15A, p55, and CASK proteins at the stereociliary tip of inner ear hair cells; and with ADGRV1, usherin, and PDZD7 at the stereociliary base in inner ear hair cells. Mutations in the gene encoding whirlin (WHRN; also known as USH2D and DFNB31), have been found to cause either USH2 subtype (USH2D) or autosomal recessive non-syndromic deafness type 31 (DFNB31). Whirlin is the key protein in the USH2 complex (whirlin, usherin and GPR98) which recruits other USH2 causative proteins at the periciliary membrane in photoreceptors and the ankle link of the stereocilia in hair cells. Whirlin's interaction with espin, another stereociliary protein, may be important for the architecture of the USH2 complex. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This whirlin family PDZ3 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F Pssm-ID: 467224 [Multi-domain] Cd Length: 91 Bit Score: 36.95 E-value: 6.21e-03
|
||||||||
| PDZ1_MAGI-1_3-like | cd06731 | PDZ domain 1 of membrane-associated guanylate kinase inverted 1 (MAGI-1), MAGI-2, and MAGI-3, ... |
441-478 | 6.28e-03 | ||||
PDZ domain 1 of membrane-associated guanylate kinase inverted 1 (MAGI-1), MAGI-2, and MAGI-3, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of MAGI1, 2, 3 (MAGI is also known as Membrane-associated guanylate kinase, WW and PDZ domain-containing protein) and related domains. MAGI proteins have been implicated in the control of cell migration and invasion through altering the activity of phosphatase and tensin homolog (PTEN) and modulating Akt signaling. Four MAGI proteins have been identified (MAGI1-3 and MAGIX). MAGI1-3 have 6 PDZ domains and bind to the C-terminus of PTEN via their PDZ2 domain. MAGIX has a single PDZ domain that is related to MAGI1-3 PDZ domain 5. Other binding partners for MAGI1 include JAM4, C-terminal tail of high risk HPV-18 E6, megalin, TRAF6, Kir4.1 (basolateral K+ channel subunit), and cadherin 23; for MAGI2, include DASM1, dendrin, axin, beta- and delta-catenin, neuroligin, hyperpolarization-activated cation channels, beta1-adrenergic receptors, NMDA receptor, and TARPs; and for MAGI3 includes LPA2. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This MAGI family PDZ1 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as beta-strands A, -B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467213 [Multi-domain] Cd Length: 85 Bit Score: 36.80 E-value: 6.28e-03
|
||||||||
| RING-HC_TRIM56_C-V | cd16584 | RING finger, HC subclass, found in tripartite motif-containing protein 56 (TRIM56) and similar ... |
16-57 | 6.80e-03 | ||||
RING finger, HC subclass, found in tripartite motif-containing protein 56 (TRIM56) and similar proteins; TRIM56, also known as RING finger protein 109 (RNF109), is a virus-inducible E3 ubiquitin ligase that restricts pestivirus infection. It positively regulates the Toll-like receptor 3 (TLR3) antiviral signaling pathway, and possesses antiviral activity against bovine viral diarrhea virus (BVDV), a ruminant pestivirus classified within the family Flaviviridae shared by tick-borne encephalitis virus (TBEV). It also possesses antiviral activity against two classical flaviviruses, yellow fever virus (YFV) and dengue virus (DENV), as well as a human coronavirus, HCoV-OC43, which is responsible for a significant share of common cold cases. It may not act on positive-strand RNA viruses indiscriminately. Moreover, TRIM56 is an interferon-inducible E3 ubiquitin ligase that modulates STING to confer double-stranded DNA-mediated innate immune responses. TRIM56 belongs to the C-V subclass of the TRIM (tripartite motif) family of proteins that are defined by an N-terminal RBCC (RING, Bbox, and coiled coil) domain, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region, as well as an uncharacterized region positioned C-terminal to the RBCC domain. Pssm-ID: 438246 [Multi-domain] Cd Length: 56 Bit Score: 36.12 E-value: 6.80e-03
|
||||||||
| RING-HC_TRIM31_C-V | cd16582 | RING finger, HC subclass, found in tripartite motif-containing protein 31 (TRIM31) and similar ... |
15-42 | 6.80e-03 | ||||
RING finger, HC subclass, found in tripartite motif-containing protein 31 (TRIM31) and similar proteins; TRIM31 is an E3 ubiquitin-protein ligase that primarily localizes to the cytoplasm, but is also associated with the mitochondria. It can negatively regulate cell proliferation and may be a potential biomarker of gastric cancer as it is overexpressed from the early stage of gastric carcinogenesis. TRIM31 is downregulated in non-small cell lung cancer and serves as a potential tumor suppressor. It interacts with p52 (Shc) and inhibits Src-induced anchorage-independent growth. TRIM31 belongs to the C-V subclass of the TRIM (tripartite motif) family of proteins that are defined by an N-terminal RBCC (RING, Bbox, and coiled coil) domain, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region, as well as an uncharacterized region positioned C-terminal to the RBCC domain. Pssm-ID: 438244 [Multi-domain] Cd Length: 44 Bit Score: 35.57 E-value: 6.80e-03
|
||||||||
| RING-HC_BARD1 | cd16496 | RING finger, HC subclass, found in BRCA1-associated RING domain protein 1 (BARD-1) and similar ... |
16-42 | 7.17e-03 | ||||
RING finger, HC subclass, found in BRCA1-associated RING domain protein 1 (BARD-1) and similar proteins; BARD-1 is a critical factor in BRCA1-mediated tumor suppression and may also serve as a target for tumorigenic lesions in some human cancers. It associates with BRCA1 (breast cancer-1) to form a heterodimeric BRCA1/BARD1 complex that is responsible for maintaining genomic stability through nuclear functions involving DNA damage signaling and repair, transcriptional regulation, and cell cycle control. The BRCA1/BARD1 complex catalyzes autoubiquitination of BRCA1 and trans ubiquitination of other protein substrates. Its E3 ligase activity is dramatically reduced in the presence of UBX domain protein 1 (UBXN1). BARD-1 contains an C3HC4-type RING-HC finger that binds BRCA1 at its N-terminus and three tandem ankyrin repeats and tandem BRCT repeat domains at its C-terminus. The BRCT repeats bind CstF-50 (cleavage stimulation factor) to modulate mRNA processing and RNAP II stability in response to DNA damage. Pssm-ID: 438159 [Multi-domain] Cd Length: 86 Bit Score: 36.54 E-value: 7.17e-03
|
||||||||
| PDZ4_GRIP1-2-like | cd06686 | PDZ domain 4 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related ... |
245-335 | 7.27e-03 | ||||
PDZ domain 4 of glutamate receptor-interacting protein 1 (GRIP1) and GRIP2, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor (AMPAR) binding proteins GRIP1 (ABP/GRIP2) and GRIP2, and related domains. GRIP1 and GRIP2 each have 7 PDZ domains. The interaction of GRIP1 and GRIP2 with GluA2/3 (AMPAR subunit) regulates AMPAR trafficking and synaptic targeting. GRIP1 has an essential role in regulating AMPAR trafficking during synaptic plasticity and learning and memory. GRIP1 and GRIP2 interact with a variety of other proteins associated with protein trafficking and internalization, for example GRIP1 also interacts with KIF5 (also known as kinesin 1), EphB receptors, scaffold protein liprin-alpha, and the rasGEF GRASP-1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This GRIP family PDZ4 domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467174 [Multi-domain] Cd Length: 99 Bit Score: 36.94 E-value: 7.27e-03
|
||||||||
| PDZ1_hSTXBP4-PDZ2_GgSTXBP4-like | cd06698 | PDZ1 domain of human syntaxin-binding protein 4 (STXBP4), PDZ2 domain of Gallus gallus ... |
256-333 | 7.56e-03 | ||||
PDZ1 domain of human syntaxin-binding protein 4 (STXBP4), PDZ2 domain of Gallus gallus uncharacterized STXBP4 isoform X1, and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of human syntaxin-binding protein 4 (STXBP4), PDZ2 domain of Gallus gallus uncharacterized STXBP4 isoform X1, and related domains. Human STXBP4 (also known as Synip) includes a single PDZ domain, a coiled-coil domain, and a WW domain (named for its two conserved tryptophans); Gallus gallus STXBP4 isoform X1 contains 2 PDZ domains (PDZ1 and PDZ2). Human STXBP4 plays a role in the translocation of transport vesicles from the cytoplasm to the plasma membrane: insulin induces the dissociation of the STXBP4 and STX4 complex liberating STX4 to interact with Vamp2, and to form the SNARE complex thereby promoting vesicle fusion. It may also play a role in the regulation of insulin release by pancreatic beta cells after stimulation by glucose. Human STXBP4 is also known to physically associate with a prominent isoform of TP63 (deltaNp63alpha 9) whose overexpression promotes squamous cell carcinoma development, and in doing so prevents degradation of this isoform by the Cdc20-APC/C complex, Itch, and RACK1. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This STXBP4-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467184 [Multi-domain] Cd Length: 89 Bit Score: 36.90 E-value: 7.56e-03
|
||||||||
| cpPDZ1_DegP-like | cd10839 | circularly permuted first PDZ domain (PDZ1) of Escherichia coli periplasmic serine ... |
446-480 | 8.29e-03 | ||||
circularly permuted first PDZ domain (PDZ1) of Escherichia coli periplasmic serine endoprotease DegP and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain 1 of Escherichia coli DegP (also known as heat shock protein DegP and Protease Do) and related domains. DegP belongs to the HtrA family of housekeeping proteases. It acts as a protease, degrading transiently denatured and unfolded or misfolded proteins which accumulate in the periplasm following heat shock or other stress conditions, and as a molecular chaperone at low temperatures. DegP has two PDZ domains in addition to the protease domain; its PDZ1 domain is responsible for identifying the distinct substrate sequences that affect degradation (degron) of the substrate sequence, and its PDZ2 domain is responsible for combining with other DegP monomers to form a stable oligomer structure. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains and as well as those with circular permutations and domain swapping of beta-strands. The canonical PDZ domain contains six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2); arranged as A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. This DegP family PDZ domain 1 is a circularly permuted PDZ domain which places beta-strand A on the C-terminus. Another permutation exists in the PDZ superfamily which places both beta-strands A and B on the C-terminus. Pssm-ID: 467630 [Multi-domain] Cd Length: 91 Bit Score: 36.69 E-value: 8.29e-03
|
||||||||
| RING-HC_Cbl | cd16708 | RING finger, HC subclass, found in E3 ubiquitin-protein ligase Cbl and similar proteins; Cbl, ... |
18-64 | 8.46e-03 | ||||
RING finger, HC subclass, found in E3 ubiquitin-protein ligase Cbl and similar proteins; Cbl, also known as Casitas B-lineage lymphoma proto-oncogene, proto-oncogene c-Cbl, RING finger protein 55 (RNF55), or signal transduction protein Cbl, is a multi-domain protein that acts as a key negative regulator of various receptor and non-receptor tyrosine kinase signaling. It contains a tyrosine kinase-binding domain (TKB, also known as the phosphotyrosine binding PTB domain, composed of a four helix-bundle, a Ca2+ binding EF-hand and a highly variant SH2 domain), a proline-rich domain, a C3HC4-type RING-HC finger, and an ubiquitin-associated (UBA) domain. TKB is responsible for the interactions with many tyrosine kinases, such as the colony-stimulating factor-1 (CSF-1) receptor, Syk/ZAP-70, and Src-family of protein tyrosine kinases. The proline-rich domain can recruit proteins with a SH3 domain. Moreover, Cbl functions as an E3 ubiquitin ligase that can bind ubiquitin-conjugating enzymes (E2s) through the RING-HC finger. Pssm-ID: 438368 [Multi-domain] Cd Length: 77 Bit Score: 36.22 E-value: 8.46e-03
|
||||||||
| RING-H2_RNF111 | cd16681 | RING finger, H2 subclass, found in RING finger protein 111 (RNF111) and similar proteins; ... |
13-63 | 8.65e-03 | ||||
RING finger, H2 subclass, found in RING finger protein 111 (RNF111) and similar proteins; RNF111, also known as Arkadia, is a nuclear E3 ubiquitin-protein ligase that targets intracellular effectors and modulators of transforming growth factor beta (TGF-beta)/Nodal-related signaling for polyubiquitination and proteasome-dependent degradation. It acts as an amplifier of Nodal signals, and enhances the dorsalizing activity of limiting amounts of Xnr1, a Nodal homolog, and requires Nodal signaling for its function. The loss of RNF111 results in early embryonic lethality, with defects attributed to compromised Nodal signaling. RNF111 also regulates tumor metastasis by modulation of the TGF-beta pathway. Its ubiquitination can be modulated by the four and a half LIM-only protein 2 (FHL2) that activates TGF-beta signal transduction. Furthermore, RNF111 interacts with the clathrin-adaptor 2 (AP2) complex and regulates endocytosis of certain cell surface receptors, leading to modulation of epidermal growth factor (EGF) and possibly other signaling pathways. In addition, RNF111 has been identified as a small ubiquitin-like modifier (SUMO)-binding protein with clustered SUMO-interacting motifs (SIMs) that together form a SUMO-binding domain (SBD). It thus functions as a SUMO-targeted ubiquitin ligase (STUbL) that directly links nonproteolytic ubiquitylation and SUMOylation in the DNA damage response, as well as triggers degradation of signal-induced polysumoylated proteins, such as the promyelocytic leukemia protein (PML). The N-terminal half of RNF111 harbors three SIMs. Its C-terminal half show high sequence similarity with RING finger protein 165 (RNF165), where it contains two serine rich domains, two nuclear localization signals, an NRG-TIER domain, and a C-terminal C3H2C3-type RING-H2 finger that is required for polyubiqutination and proteasome-dependent degradation of phosphorylated forms of Smad2/3 and three major negative regulators of TGF-beta signaling, Smad7, SnoN and c-Ski. Pssm-ID: 438343 [Multi-domain] Cd Length: 61 Bit Score: 35.81 E-value: 8.65e-03
|
||||||||
| PDZ_ZASP52-like | cd23068 | PDZ domain of Drosophila melanogaster PDZ and LIM domain protein Zasp52 (also known as Zasp), ... |
447-502 | 8.73e-03 | ||||
PDZ domain of Drosophila melanogaster PDZ and LIM domain protein Zasp52 (also known as Zasp), and related domains; PDZ (PSD-95 (Postsynaptic density protein 95), Dlg (Discs large protein), and ZO-1 (Zonula occludens-1)) domain of Drosophila melanogaster Zasp52 and related domains. Drosophila melanogaster Zasp52 (also known as Z band alternatively spliced PDZ-motif protein or Zasp) colocalizes with integrins at myotendinous junctions and with alpha-actinin at Z-disks and is required for muscle attachment as well as Z-disk assembly and maintenance. The Zasp52 actin-binding site includes the extended PDZ domain and the ZM region. The Zasp52-PDZ domain is required for myofibril assembly. PDZ domains usually bind in a sequence-specific manner to short peptide sequences located at the C-terminal end of their partner proteins (known as PDZ binding motifs). The PDZ superfamily includes canonical PDZ domains as well as those with circular permutations and domain swapping mediated by beta-strands. This Zasp52-like family domain is a canonical PDZ domain containing six beta-strands A-F and two alpha-helices (alpha-helix 1 and 2), arranged in the order: beta-strands A, B, C, alpha-helix 1, beta-strands D, E, alpha-helix 2 and beta-strand F. Pssm-ID: 467281 [Multi-domain] Cd Length: 82 Bit Score: 36.35 E-value: 8.73e-03
|
||||||||
| RING-HC_CeBARD1-like | cd23143 | RING finger, HC subclass, found in Caenorhabditis elegans BRCA1-associated RING domain protein ... |
16-61 | 8.93e-03 | ||||
RING finger, HC subclass, found in Caenorhabditis elegans BRCA1-associated RING domain protein 1 (CeBARD1) and similar proteins; CeBARD1, also called Ce-BRD-1, Cebrd-1, or RING-type E3 ubiquitin transferase BARD1, is a constituent of the CeBCD complex that possesses E3 ubiquitin-protein ligase activity. It plays a role in triggering cellular responses at damage sites in response to DNA damage that may be induced by ionizing radiation. It protects against chromosome non-disjunction and nuclear fragmentation during meiotic double-strand break repair to ensure sister chromatid recombination and aid chromosome stability. CeBARD1 contains a typical C3HC4-type RING-HC finger. Pssm-ID: 438505 [Multi-domain] Cd Length: 47 Bit Score: 35.21 E-value: 8.93e-03
|
||||||||
| PLN03208 | PLN03208 | E3 ubiquitin-protein ligase RMA2; Provisional |
11-45 | 8.94e-03 | ||||
E3 ubiquitin-protein ligase RMA2; Provisional Pssm-ID: 178747 [Multi-domain] Cd Length: 193 Bit Score: 38.53 E-value: 8.94e-03
|
||||||||
| RING-HC_MID_C-I | cd16575 | RING finger, HC subclass, found in midline-1 (MID1), midline-2 (MID2) and similar proteins; ... |
16-41 | 9.24e-03 | ||||
RING finger, HC subclass, found in midline-1 (MID1), midline-2 (MID2) and similar proteins; MID1, also known as midin, midline 1 RING finger protein, putative transcription factor XPRF, RING finger protein 59 (RNF59), or tripartite motif-containing protein 18 (TRIM18), is a microtubule-associated E3 ubiquitin-protein ligase implicated in epithelial-mesenchymal differentiation, cell migration and adhesion, and programmed cell death along specific regions of the ventral midline during embryogenesis. MID2, also known as midin-2, midline defect 2, RING finger protein 60 (RNF60), or tripartite motif-containing protein 1 (TRIM1), associates with the microtubule network and may at least partially compensate for the loss of MID1. Both MID1 and MID2 interacts with Alpha 4, which is a regulatory subunit of PP2-type phosphatases, such as PP2A, and an integral component of the rapamycin-sensitive signaling pathway. They also play a central role in the regulation of granule exocytosis. Functional redundancy exists between MID1 and MID2 in cytotoxic lymphocytes (CTL). Both MID1 and MID2 belong to the C-I subclass of the TRIM (tripartite motif) family of proteins that are defined by their N-terminal RBCC (RING, Bbox, and coiled coil) domains, including three consecutive zinc-binding domains, a C3HC4-type RING-HC finger, Bbox1 and Bbox2, and a coiled coil region, as well as a COS (carboxyl-terminal subgroup one signature) box, a fibronectin type III (FN3) domain, and a B30.2/SPRY (SplA and ryanodine receptor) domain positioned C-terminal to the RBCC domain. Pssm-ID: 438237 [Multi-domain] Cd Length: 54 Bit Score: 35.67 E-value: 9.24e-03
|
||||||||
| RING-H2_MBR | cd23113 | RING finger, H2 subclass, found in Arabidopsis thaliana MED25-binding RING-H2 protein (MBR) ... |
15-53 | 9.46e-03 | ||||
RING finger, H2 subclass, found in Arabidopsis thaliana MED25-binding RING-H2 protein (MBR) and similar proteins; This subfamily includes MBR1 and MBR2 (also called HAL3-interacting protein 1 or AtHIP1). They are E3 ubiquitin-protein ligases that function as regulators of MED25 stability by targeting MED25 for degradation in a RING-H2-dependent manner. Proteasome-dependent degradation of MED25 seems to activate its function as a positive regulator of FLOWERING LOCUS T (FT) and is important to induce the expression of FT, and consequently to promote flowering. MBR2 may also function downstream of HAL3 and be required for HAL3-regulated plant growth. Activation of MBR2 by HAL3 may lead to the degradation of cell cycle suppressors, resulting in enhancement of cell division and plant growth. Both MBR1 and MBR2 contain a C3H2C3-type RING-H2 finger. Pssm-ID: 438475 [Multi-domain] Cd Length: 50 Bit Score: 35.24 E-value: 9.46e-03
|
||||||||
| RING-HC_RNF219 | cd16562 | RING finger, HC subclass, found in RING finger protein 219 (RNF219) and similar proteins; ... |
16-57 | 9.73e-03 | ||||
RING finger, HC subclass, found in RING finger protein 219 (RNF219) and similar proteins; RNF219 may function as a modulator of late-onset Alzheimer's disease (LOAD) associated amyloid beta A4 precursor protein (APP) endocytosis and metabolism. It genetically interacts with apolipoprotein E epsilon4 allele (APOE4). Thus, a genetic variant of RNF219 was found to affect amyloid deposition in human brain and LOAD age-of-onset. Moreover, common genetic variants at the RNF219 locus had been associated with alternations in lipid metabolism, cognitive performance and central nervous system ventricle volume. RNF219 contains a C3HC4-type RING-HC finger. Pssm-ID: 438224 [Multi-domain] Cd Length: 45 Bit Score: 35.10 E-value: 9.73e-03
|
||||||||
| RING-HC_RNF170 | cd16553 | RING finger, HC subclass, found in RING finger protein 170 (RNF170) and similar proteins; ... |
15-57 | 9.90e-03 | ||||
RING finger, HC subclass, found in RING finger protein 170 (RNF170) and similar proteins; RNF170, also known as putative LAG1-interacting protein, is an endoplasmic reticulum (ER) membrane-bound E3 ubiquitin-protein ligase that mediates ubiquitination-dependent degradation of type-I inositol 1,4,5-trisphosphate (IP3) receptors (ITPR1) via the endoplasmic-reticulum-associated protein degradation (ERAD) pathway. A point mutation (arginine to cysteine at position 199) in the RNF170 gene is linked with autosomal-dominant sensory ataxia (ADSA), a disease characterized by neurodegeneration in the posterior columns of the spinal cord. RNF170 contains a C3HC4-type RING-HC finger. Pssm-ID: 438215 [Multi-domain] Cd Length: 57 Bit Score: 35.34 E-value: 9.90e-03
|
||||||||
| Blast search parameters | ||||
|
