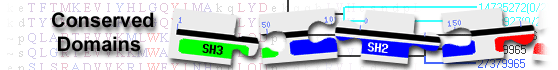


1-phosphatidylinositol 4,5-bisphosphate phosphodiesterase zeta-1 isoform X4 [Mus musculus]
Protein Classification

PLC family C2 domain-containing protein( domain architecture ID 11979330)
PLC (phosphoinositide-specific phospholipase C) family C2 domain-containing protein similar to C2 domain region of PLCs that are involved in the hydrolysis of phosphatidylinositol-4,5-bisphosphate (PIP2) to d-myo-inositol-1,4,5-trisphosphate (1,4,5-IP3) and sn-1,2-diacylglycerol (DAG)
List of domain hits
| Name | Accession | Description | Interval | E-value | ||||||
| PI-PLCc_GDPD_SF super family | cl14615 | Catalytic domain of phosphoinositide-specific phospholipase C-like phosphodiesterases ... |
52-379 | 2.91e-163 | ||||||
Catalytic domain of phosphoinositide-specific phospholipase C-like phosphodiesterases superfamily; The PI-PLC-like phosphodiesterases superfamily represents the catalytic domains of bacterial phosphatidylinositol-specific phospholipase C (PI-PLC, EC 4.6.1.13), eukaryotic phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11), glycerophosphodiester phosphodiesterases (GP-GDE, EC 3.1.4.46), sphingomyelinases D (SMases D) (sphingomyelin phosphodiesterase D, EC 3.1.4.41) from spider venom, SMases D-like proteins, and phospholipase D (PLD) from several pathogenic bacteria, as well as their uncharacterized homologs found in organisms ranging from bacteria and archaea to metazoans, plants, and fungi. PI-PLCs are ubiquitous enzymes hydrolyzing the membrane lipid phosphoinositides to yield two important second messengers, inositol phosphates and diacylglycerol (DAG). GP-GDEs play essential roles in glycerol metabolism and catalyze the hydrolysis of glycerophosphodiesters to sn-glycerol-3-phosphate (G3P) and the corresponding alcohols that are major sources of carbon and phosphate. Both, PI-PLCs and GP-GDEs, can hydrolyze the 3'-5' phosphodiester bonds in different substrates, and utilize a similar mechanism of general base and acid catalysis with conserved histidine residues, which consists of two steps, a phosphotransfer and a phosphodiesterase reaction. This superfamily also includes Neurospora crassa ankyrin repeat protein NUC-2 and its Saccharomyces cerevisiae counterpart, Phosphate system positive regulatory protein PHO81, glycerophosphodiester phosphodiesterase (GP-GDE)-like protein SHV3 and SHV3-like proteins (SVLs). The residues essential for enzyme activities and metal binding are not conserved in these sequence homologs, which might suggest that the function of catalytic domains in these proteins might be distinct from those in typical PLC-like phosphodiesterases. The actual alignment was detected with superfamily member cd08595: Pssm-ID: 472694 [Multi-domain] Cd Length: 257 Bit Score: 465.57 E-value: 2.91e-163
|
||||||||||
| C2_PLC_like | cd00275 | C2 domain present in Phosphoinositide-specific phospholipases C (PLC); PLCs are involved in ... |
409-527 | 3.73e-32 | ||||||
C2 domain present in Phosphoinositide-specific phospholipases C (PLC); PLCs are involved in the hydrolysis of phosphatidylinositol-4,5-bisphosphate (PIP2) to d-myo-inositol-1,4,5-trisphosphate (1,4,5-IP3) and sn-1,2-diacylglycerol (DAG). 1,4,5-IP3 and DAG are second messengers in eukaryotic signal transduction cascades. PLC is composed of a N-terminal PH domain followed by a series of EF hands, a catalytic TIM barrel and a C-terminal C2 domain. C2 domains fold into an 8-standed beta-sandwich that can adopt 2 structural arrangements: Type I and Type II, distinguished by a circular permutation involving their N- and C-terminal beta strands. Many C2 domains are Ca2+-dependent membrane-targeting modules that bind a wide variety of substances including bind phospholipids, inositol polyphosphates, and intracellular proteins. Most C2 domain proteins are either signal transduction enzymes that contain a single C2 domain, such as protein kinase C, or membrane trafficking proteins which contain at least two C2 domains, such as synaptotagmin 1. However, there are a few exceptions to this including RIM isoforms and some splice variants of piccolo/aczonin and intersectin which only have a single C2 domain. C2 domains with a calcium binding region have negatively charged residues, primarily aspartates, that serve as ligands for calcium ions. Members here have a type-II topology. : Pssm-ID: 175974 [Multi-domain] Cd Length: 128 Bit Score: 120.34 E-value: 3.73e-32
|
||||||||||
| EFh_PI-PLC super family | cl28895 | EF-hand motif found in eukaryotic phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4. ... |
2-40 | 2.36e-08 | ||||||
EF-hand motif found in eukaryotic phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11) isozymes; PI-PLC isozymes are signaling enzymes that hydrolyze the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, Inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which goes on to phosphorylate other molecules, leading to altered cellular activity. Calcium is required for the catalysis. This family corresponds to the four EF-hand motifs containing PI-PLC isozymes, including PI-PLC-beta (1-4), -gamma (1-2), -delta (1,3,4), -epsilon (1), -zeta (1), eta (1-2). Lower eukaryotes such as yeast and slime molds contain only delta-type isozymes. In contrast, other types of isoforms present in higher eukaryotes. This family also includes 1-phosphatidylinositol 4,5-bisphosphate phosphodiesterase 1 (PLC1) from fungi. Some homologs from plants contain only two atypical EF-hand motifs and they are not included. All PI-PLC isozymes except sperm-specific PI-PLC-zeta share a core set of domains, including an N-terminal pleckstrin homology (PH) domain, four atypical EF-hand motifs, a PLC catalytic core, and a single C2 domain. PI-PLC-zeta lacks the PH domain. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. Most of EF-hand motifs found in PI-PLCs consist of a helix-loop-helix structure, but lack residues critical to metal binding. Moreover, the EF-hand region of most of PI-PLCs may have an important regulatory function, but it has yet to be identified. However, PI-PLC-zeta is a key exception. It is responsible for Ca2+ oscillations in fertilized oocytes and exhibits a high sensitivity to Ca2+ mediated through its EF-hand domain. In addition, PI-PLC-eta2 shows a canonical EF-loop directing Ca2+-sensitivity and thus can amplify transient Ca2+ signals. Also it appears that PI-PLC-delta1 can regulate the binding of PH domain to PIP2 in a Ca2+-dependent manner through its functionally important EF-hand domains. PI-PLCs can be activated by a variety of extracellular ligands, such as growth factors, hormones, cytokines and lipids. Their activation has been implicated in tumorigenesis and/or metastasis linked to migration, proliferation, growth, inflammation, angiogenesis and actin cytoskeleton reorganization. PI-PLC-beta isozymes are activated by G-protein coupled receptor (GPCR) through different mechanisms. However, PI-PLC-gamma isozymes are activated by receptor tyrosine kinase (RTK), such as Rho and Ras GTPases. In contrast, PI-PLC-epsilon are activated by both GPCR and RTK. PI-PLC-delta1 and PLC-eta 1 are activated by GPCR-mediated calcium mobilization. The activation mechanism for PI-PLC-zeta remains unclear. The actual alignment was detected with superfamily member cd16204: Pssm-ID: 333715 [Multi-domain] Cd Length: 142 Bit Score: 53.28 E-value: 2.36e-08
|
||||||||||
| Name | Accession | Description | Interval | E-value | |||||||
| PI-PLCc_zeta | cd08595 | Catalytic domain of metazoan phosphoinositide-specific phospholipase C-zeta; This family ... |
52-379 | 2.91e-163 | |||||||
Catalytic domain of metazoan phosphoinositide-specific phospholipase C-zeta; This family corresponds to the catalytic domain presenting in metazoan phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11)-zeta isozyme. PI-PLC is a signaling enzyme that hydrolyzes the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which then phosphorylates other molecules, leading to altered cellular activity. Calcium is required for the catalysis. PI-PLC-zeta represents a class of sperm-specific PI-PLC that has an N-terminal EF-hand domain, a PLC catalytic core domain, and a C-terminal C2 domain. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. There is one PLC-zeta isozyme (1). PLC-zeta plays a fundamental role in vertebrate fertilization by initiating intracellular calcium oscillations that trigger the embryo development. However, the mechanism of its activation still remains unclear. Aside from PI-PLC-zeta identified in mammals, its eukaryotic homologs have been classified with this family. Pssm-ID: 176537 [Multi-domain] Cd Length: 257 Bit Score: 465.57 E-value: 2.91e-163
|
|||||||||||
| PI-PLC-X | pfam00388 | Phosphatidylinositol-specific phospholipase C, X domain; This associates with pfam00387 to ... |
55-196 | 9.24e-81 | |||||||
Phosphatidylinositol-specific phospholipase C, X domain; This associates with pfam00387 to form a single structural unit. Pssm-ID: 459795 [Multi-domain] Cd Length: 142 Bit Score: 249.73 E-value: 9.24e-81
|
|||||||||||
| PLN02952 | PLN02952 | phosphoinositide phospholipase C |
51-504 | 4.79e-67 | |||||||
phosphoinositide phospholipase C Pssm-ID: 178538 [Multi-domain] Cd Length: 599 Bit Score: 228.73 E-value: 4.79e-67
|
|||||||||||
| PLCXc | smart00148 | Phospholipase C, catalytic domain (part); domain X; Phosphoinositide-specific phospholipases C. ... |
55-197 | 5.12e-61 | |||||||
Phospholipase C, catalytic domain (part); domain X; Phosphoinositide-specific phospholipases C. These enzymes contain 2 regions (X and Y) which together form a TIM barrel-like structure containing the active site residues. Phospholipase C enzymes (PI-PLC) act as signal transducers that generate two second messengers, inositol-1,4,5-trisphosphate and diacylglycerol. The bacterial enzyme appears to be a homologue of the mammalian PLCs. Pssm-ID: 197543 [Multi-domain] Cd Length: 143 Bit Score: 198.27 E-value: 5.12e-61
|
|||||||||||
| C2_PLC_like | cd00275 | C2 domain present in Phosphoinositide-specific phospholipases C (PLC); PLCs are involved in ... |
409-527 | 3.73e-32 | |||||||
C2 domain present in Phosphoinositide-specific phospholipases C (PLC); PLCs are involved in the hydrolysis of phosphatidylinositol-4,5-bisphosphate (PIP2) to d-myo-inositol-1,4,5-trisphosphate (1,4,5-IP3) and sn-1,2-diacylglycerol (DAG). 1,4,5-IP3 and DAG are second messengers in eukaryotic signal transduction cascades. PLC is composed of a N-terminal PH domain followed by a series of EF hands, a catalytic TIM barrel and a C-terminal C2 domain. C2 domains fold into an 8-standed beta-sandwich that can adopt 2 structural arrangements: Type I and Type II, distinguished by a circular permutation involving their N- and C-terminal beta strands. Many C2 domains are Ca2+-dependent membrane-targeting modules that bind a wide variety of substances including bind phospholipids, inositol polyphosphates, and intracellular proteins. Most C2 domain proteins are either signal transduction enzymes that contain a single C2 domain, such as protein kinase C, or membrane trafficking proteins which contain at least two C2 domains, such as synaptotagmin 1. However, there are a few exceptions to this including RIM isoforms and some splice variants of piccolo/aczonin and intersectin which only have a single C2 domain. C2 domains with a calcium binding region have negatively charged residues, primarily aspartates, that serve as ligands for calcium ions. Members here have a type-II topology. Pssm-ID: 175974 [Multi-domain] Cd Length: 128 Bit Score: 120.34 E-value: 3.73e-32
|
|||||||||||
| C2 | smart00239 | Protein kinase C conserved region 2 (CalB); Ca2+-binding motif present in phospholipases, ... |
411-514 | 5.79e-18 | |||||||
Protein kinase C conserved region 2 (CalB); Ca2+-binding motif present in phospholipases, protein kinases C, and synaptotagmins (among others). Some do not appear to contain Ca2+-binding sites. Particular C2s appear to bind phospholipids, inositol polyphosphates, and intracellular proteins. Unusual occurrence in perforin. Synaptotagmin and PLC C2s are permuted in sequence with respect to N- and C-terminal beta strands. SMART detects C2 domains using one or both of two profiles. Pssm-ID: 214577 [Multi-domain] Cd Length: 101 Bit Score: 79.45 E-value: 5.79e-18
|
|||||||||||
| C2 | pfam00168 | C2 domain; |
411-503 | 1.56e-15 | |||||||
C2 domain; Pssm-ID: 425499 [Multi-domain] Cd Length: 104 Bit Score: 72.35 E-value: 1.56e-15
|
|||||||||||
| EFh_PI-PLCzeta | cd16204 | EF-hand motif found in phosphoinositide phospholipase C zeta 1 (PI-PLC-zeta1); PI-PLC-zeta1, ... |
2-40 | 2.36e-08 | |||||||
EF-hand motif found in phosphoinositide phospholipase C zeta 1 (PI-PLC-zeta1); PI-PLC-zeta1, also termed 1-phosphatidylinositol 4,5-bisphosphate phosphodiesterase zeta-1, or phospholipase C-zeta-1 (PLC-zeta-1), or testis-development protein NYD-SP27, is only found in the testis. The sperm-specific PI-PLC plays a fundamental role in vertebrate fertilization by initiating intracellular calcium oscillations that trigger the embryo development. However, the mechanism of its activation still remains unclear. PI-PLC-zeta1 contains an N-terminal four atypical EF-hand motifs, a PLC catalytic core domain, and a C-terminal C2 domain. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. Unlike other PI-PLCs, PI-PLC-zeta is responsible for Ca2+ oscillations in fertilized oocytes and exhibits a high sensitivity to Ca2+ mediated through its EF-hand domain. There is only one PLC-zeta isozyme. Aside from PI-PLC-zeta identified in mammals, its eukaryotic homologs have been classified with this family. Pssm-ID: 320034 [Multi-domain] Cd Length: 142 Bit Score: 53.28 E-value: 2.36e-08
|
|||||||||||
| EF-hand_like | pfam09279 | Phosphoinositide-specific phospholipase C, efhand-like; Members of this family are ... |
1-44 | 1.72e-05 | |||||||
Phosphoinositide-specific phospholipase C, efhand-like; Members of this family are predominantly found in phosphoinositide-specific phospholipase C. They adopt a structure consisting of a core of four alpha helices, in an EF like fold, and are required for functioning of the enzyme. Pssm-ID: 401279 [Multi-domain] Cd Length: 85 Bit Score: 43.39 E-value: 1.72e-05
|
|||||||||||
| Name | Accession | Description | Interval | E-value | ||||||||
| PI-PLCc_zeta | cd08595 | Catalytic domain of metazoan phosphoinositide-specific phospholipase C-zeta; This family ... |
52-379 | 2.91e-163 | ||||||||
Catalytic domain of metazoan phosphoinositide-specific phospholipase C-zeta; This family corresponds to the catalytic domain presenting in metazoan phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11)-zeta isozyme. PI-PLC is a signaling enzyme that hydrolyzes the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which then phosphorylates other molecules, leading to altered cellular activity. Calcium is required for the catalysis. PI-PLC-zeta represents a class of sperm-specific PI-PLC that has an N-terminal EF-hand domain, a PLC catalytic core domain, and a C-terminal C2 domain. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. There is one PLC-zeta isozyme (1). PLC-zeta plays a fundamental role in vertebrate fertilization by initiating intracellular calcium oscillations that trigger the embryo development. However, the mechanism of its activation still remains unclear. Aside from PI-PLC-zeta identified in mammals, its eukaryotic homologs have been classified with this family. Pssm-ID: 176537 [Multi-domain] Cd Length: 257 Bit Score: 465.57 E-value: 2.91e-163
|
||||||||||||
| PI-PLCc_delta | cd08593 | Catalytic domain of metazoan phosphoinositide-specific phospholipase C-delta; This subfamily ... |
52-379 | 3.72e-129 | ||||||||
Catalytic domain of metazoan phosphoinositide-specific phospholipase C-delta; This subfamily corresponds to the catalytic domain present in metazoan phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11)-delta isozymes. PI-PLC is a signaling enzyme that hydrolyzes the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, Inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which then phosphorylates other molecules, leading to altered cellular activity. Calcium is required for the catalysis. PLC-delta represents a class of mammalian PI-PLC that has an N-terminal pleckstrin homology (PH) domain, an array of EF hands, a PLC catalytic core domain, and a C-terminal C2 domain. This CD corresponds to the catalytic domain which is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. There are three PI-PLC-delta isozymes (1,3 and 4). PI-PLC-delta1 is relatively well characterized. It is activated by high calcium levels generated by other PI-PLC family members, and therefore functions as a calcium amplifier within the cell. Different PI-PLC-delta isozymes have different tissue distribution and different subcellular locations. PI-PLC-delta1 is mostly a cytoplasmic protein, PI-PLC-delta3 is located in the membrane, and PI-PLC-delta4 is predominantly detected in the cell nucleus. Aside from three PI-PLC-delta isozymes identified in mammals, some eukaryotic PI-PLC-delta homologs have been classified to this CD. Pssm-ID: 176535 [Multi-domain] Cd Length: 257 Bit Score: 378.60 E-value: 3.72e-129
|
||||||||||||
| PI-PLCc_eukaryota | cd08558 | Catalytic domain of eukaryotic phosphoinositide-specific phospholipase C and similar proteins; ... |
52-379 | 5.75e-127 | ||||||||
Catalytic domain of eukaryotic phosphoinositide-specific phospholipase C and similar proteins; This family corresponds to the catalytic domain present in eukaryotic phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11) and similar proteins. The higher eukaryotic PI-PLCs play a critical role in most signal transduction pathways, controlling numerous cellular events such as cell growth, proliferation, excitation and secretion. They strictly require Ca2+ for the catalytic activity. They display a clear preference towards the hydrolysis of the more highly phosphorylated membrane phospholipids PI-analogues, phosphatidylinositol 4,5-bisphosphate (PIP2) and phosphatidylinositol-4-phosphate (PIP), to generate two important second messengers in eukaryotic signal transduction cascades, inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which then phosphorylates other molecules, leading to altered cellular activity. The eukaryotic PI-PLCs have a multidomain organization that consists of a PLC catalytic core domain, and various regulatory domains, such as the pleckstrin homology (PH) domain, EF-hand motif, and C2 domain. The catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a linker region. The catalytic mechanism of eukaryotic PI-PLCs is based on general base and acid catalysis utilizing two well conserved histidines and consists of two steps, a phosphotransfer and a phosphodiesterase reaction. The mammalian PI-PLCs consist of 13 isozymes, which are classified into six-subfamilies, PI-PLC-delta (1,3 and 4), -beta(1-4), -gamma(1,2), -epsilon, -zeta, and -eta (1,2). Ca2+ is required for the activation of all forms of mammalian PI-PLCs, and the concentration of calcium influences substrate specificity. This family also includes metazoan phospholipase C related but catalytically inactive proteins (PRIP), which belong to a group of novel inositol trisphosphate binding proteins. Due to the replacement of critical catalytic residues, PRIP does not have PLC enzymatic activity. Pssm-ID: 176501 [Multi-domain] Cd Length: 226 Bit Score: 371.40 E-value: 5.75e-127
|
||||||||||||
| PI-PLCc_PRIP_metazoa | cd08597 | Catalytic domain of metazoan phospholipase C related, but catalytically inactive protein; This ... |
53-379 | 8.16e-114 | ||||||||
Catalytic domain of metazoan phospholipase C related, but catalytically inactive protein; This family corresponds to the catalytic domain present in metazoan phospholipase C related, but catalytically inactive proteins (PRIP), which belong to a group of novel Inositol 1,4,5-trisphosphate (InsP3) binding protein. PRIP has a primary structure and domain architecture, incorporating a pleckstrin homology (PH) domain, an array of EF hands, a PLC catalytic core domain with highly conserved X- and Y-regions split by a linker sequence, and a C-terminal C2 domain, similar to phosphoinositide-specific phospholipases C (PI-PLC, EC 3.1.4.11)-delta isoforms. Due to replacement of critical catalytic residues, PRIP do not have PLC enzymatic activity. PRIP consists of two subfamilies, PRIP-1(previously known as p130 or PLC-1), which is predominantly expressed in the brain, and PRIP-2 (previously known as PLC-2), which exhibits a relatively ubiquitous expression. Experiments show both, PRIP-1 and PRIP-2, are involved in InsP3-mediated calcium signaling pathway and GABA(A)receptor-mediated signaling pathway. In addition, PRIP-2 acts as a negative regulator of B-cell receptor signaling and immune responses. Pssm-ID: 176539 [Multi-domain] Cd Length: 260 Bit Score: 339.40 E-value: 8.16e-114
|
||||||||||||
| PI-PLCc_delta4 | cd08631 | Catalytic domain of metazoan phosphoinositide-specific phospholipase C-delta4; This subfamily ... |
52-379 | 5.03e-108 | ||||||||
Catalytic domain of metazoan phosphoinositide-specific phospholipase C-delta4; This subfamily corresponds to the catalytic domain present in metazoan phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11)-delta4 isozymes. PI-PLC is a signaling enzyme that hydrolyzes the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, Inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which then phosphorylates other molecules, leading to altered cellular activity. Calcium is required for the catalysis. PLC-delta represents a class of mammalian PI-PLC that has an N-terminal pleckstrin homology (PH) domain, an array of EF hands, a PLC catalytic core domain, and a C-terminal C2 domain. This CD corresponds to the catalytic domain which is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. There are three PI-PLC-delta isozymes (1,3 and 4). Unlike PI-PLC-delta 1 and 3, a putative nuclear export sequence (NES) located in the EF-hand domain, which may be responsible transporting PI-PLC-delta1 and 3 from the cell nucleus, is not present in PI-PLC-delta4. Experiments show PI-PLC-delta4 is required for the acrosome reaction in fertilization. Pssm-ID: 176568 [Multi-domain] Cd Length: 258 Bit Score: 324.59 E-value: 5.03e-108
|
||||||||||||
| PI-PLCc_delta1 | cd08629 | Catalytic domain of metazoan phosphoinositide-specific phospholipase C-delta1; This subfamily ... |
52-379 | 6.80e-102 | ||||||||
Catalytic domain of metazoan phosphoinositide-specific phospholipase C-delta1; This subfamily corresponds to the catalytic domain present in metazoan phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11)-delta1 isozymes. PI-PLC is a signaling enzyme that hydrolyzes the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, Inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which then phosphorylates other molecules, leading to altered cellular activity. Calcium is required for the catalysis. PLC-delta represents a class of mammalian PI-PLC that has an N-terminal pleckstrin homology (PH) domain, an array of EF hands, a PLC catalytic core domain, and a C-terminal C2 domain. This subfamily corresponds to the catalytic domain which is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. There are three PI-PLC-delta isozymes (1,3 and 4). PI-PLC-delta1 is relatively well characterized. It is activated by high calcium levels generated by other PI-PLC family members, and therefore functions as a calcium amplifier within the cell. Unlike PI-PLC-delta 4, PI-PLC-delta1 and 3 possess a putative nuclear export sequence (NES) located in the EF-hand domain, which may be responsible transporting PI-PLC-delta1and 3 from the cell nucleus. Experiments show PI-PLC-delta1 is essential for normal hair formation. Pssm-ID: 176566 [Multi-domain] Cd Length: 258 Bit Score: 308.89 E-value: 6.80e-102
|
||||||||||||
| PI-PLCc_delta3 | cd08630 | Catalytic domain of metazoan phosphoinositide-specific phospholipase C-delta3; This subfamily ... |
52-379 | 2.75e-97 | ||||||||
Catalytic domain of metazoan phosphoinositide-specific phospholipase C-delta3; This subfamily corresponds to the catalytic domain present in metazoan phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11)-delta3 isozymes. PI-PLC is a signaling enzyme that hydrolyzes the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, Inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which then phosphorylates other molecules, leading to altered cellular activity. Calcium is required for the catalysis. PLC-delta represents a class of mammalian PI-PLC that has an N-terminal pleckstrin homology (PH) domain, an array of EF hands, a PLC catalytic core domain, and a C-terminal C2 domain. This family corresponds to the catalytic domain which is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. There are three PI-PLC-delta isozymes (1,3 and 4). Unlike PI-PLC-delta 4, PI-PLC-delta1 and 3 possess a putative nuclear export sequence (NES) located in the EF-hand domain, which may be responsible transporting PI-PLC-delta1 and 3 from the cell nucleus. Pssm-ID: 176567 [Multi-domain] Cd Length: 258 Bit Score: 296.93 E-value: 2.75e-97
|
||||||||||||
| PI-PLC1c_yeast | cd08598 | Catalytic domain of putative yeast phosphatidylinositide-specific phospholipases C; This ... |
52-376 | 2.10e-96 | ||||||||
Catalytic domain of putative yeast phosphatidylinositide-specific phospholipases C; This family corresponds to the catalytic domain present in a group of putative phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11) encoded by PLC1 genes from yeasts, which are homologs of the delta isoforms of mammalian PI-PLC in terms of overall sequence similarity and domain organization. Mammalian PI-PLC is a signaling enzyme that hydrolyzes the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which then phosphorylates other molecules, leading to altered cellular activity. Calcium is required for the catalysis. The prototype of this CD is protein Plc1p encoded by PLC1 genes from Saccharomyces cerevisiae. Plc1p contains both highly conserved X- and Y- regions of PLC catalytic core domain, as well as a presumptive EF-hand like calcium binding motif. Experiments show that Plc1p displays calcium dependent catalytic properties with high similarity to those of the mammalian PLCs, and plays multiple roles in modulating the membrane/protein interactions in filamentation control. CaPlc1p encoded by CAPLC1 from the closely related yeast Candida albicans, an orthologue of S. cerevisiae Plc1p, is also included in this group. Like Plc1p, CaPlc1p has conserved presumptive catalytic domain, shows PLC activity when expressed in E. coli, and is involved in multiple cellular processes. There are two other gene copies of CAPLC1 in C. albicans, CAPLC2 (also named as PIPLC) and CAPLC3. Experiments show CaPlc1p is the only enzyme in C. albicans which functions as PLC. The biological functions of CAPLC2 and CAPLC3 gene products must be clearly different from CaPlc1p, but their exact roles remain unclear. Moreover, CAPLC2 and CAPLC3 gene products are more similar to extracellular bacterial PI-PLC than to the eukaryotic PI-PLC, and they are not included in this subfamily. Pssm-ID: 176540 [Multi-domain] Cd Length: 231 Bit Score: 293.38 E-value: 2.10e-96
|
||||||||||||
| PI-PLCc_beta | cd08591 | Catalytic domain of metazoan phosphoinositide-specific phospholipase C-beta; This subfamily ... |
52-376 | 4.67e-95 | ||||||||
Catalytic domain of metazoan phosphoinositide-specific phospholipase C-beta; This subfamily corresponds to the catalytic domain present in metazoan phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11)-beta isozymes. PI-PLC is a signaling enzyme that hydrolyzes the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, Inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which goes on to phosphorylate other molecules, leading to altered cellular activity. Calcium is required for the catalysis. PLC-beta represents a class of mammalian PI-PLC that has an N-terminal pleckstrin homology (PH) domain, an array of EF hands, a PLC catalytic core domain, a C2 domain, and a unique C-terminal coiled-coil (CT) domain necessary for homodimerization. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. There are four PLC-beta isozymes (1-4). They are activated by the heterotrimeric G protein alpha q subunits through their C2 domain and long C-terminal extension. The beta-gamma subunits of heterotrimeric G proteins are known to activate the PLC-beta2 and -beta3 isozymes only. Aside from four PLC-beta isozymes identified in mammals, some eukaryotic PLC-beta homologs have been classified into this subfamily, such as NorpA and PLC-21 from Drosophila and PLC-beta from turkey, Xenopus, sponge, and hydra. Pssm-ID: 176533 [Multi-domain] Cd Length: 257 Bit Score: 291.16 E-value: 4.67e-95
|
||||||||||||
| PI-PLCc_gamma | cd08592 | Catalytic domain of metazoan phosphoinositide-specific phospholipase C-gamma; This family ... |
53-379 | 5.42e-89 | ||||||||
Catalytic domain of metazoan phosphoinositide-specific phospholipase C-gamma; This family corresponds to the catalytic domain present in metazoan phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11)-gamma isozymes. PI-PLC is a signaling enzyme that hydrolyzes the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which goes on to phosphorylate other molecules, leading to altered cellular activity. Calcium is required for the catalysis. PI-PLC-gamma represents a class of mammalian PI-PLC that has an N-terminal pleckstrin homology (PH) domain, an array of EF hands, a PLC catalytic core domain, and a C2 domain.The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. Unique to PI-PLC-gamma, a second PH domain, two SH2 (Src homology 2) regions, and one SH3 (Src homology 3) region is present within this linker region. There are two PI-PLC-gamma isozymes (1-2). They are activated by receptor and non-receptor tyrosine kinases due to the presence of two SH2 and a single SH3 domain within the linker region. Aside from the two PI-PLC-gamma isozymes identified in mammals, some eukaryotic PI-PLC-gamma homologs have been classified with this subfamily. Pssm-ID: 176534 [Multi-domain] Cd Length: 229 Bit Score: 274.30 E-value: 5.42e-89
|
||||||||||||
| PI-PLCc_eta2 | cd08633 | Catalytic domain of metazoan phosphoinositide-specific phospholipase C-eta2; This subfamily ... |
53-379 | 1.93e-82 | ||||||||
Catalytic domain of metazoan phosphoinositide-specific phospholipase C-eta2; This subfamily corresponds to the catalytic domain present in metazoan phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11)-eta isozyme 2. PI-PLC is a signaling enzyme that hydrolyzes the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, Inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which then phosphorylates other molecules, leading to altered cellular activity. Calcium is required for the catalysis. PI-PLC-eta represents a class of neuron-speific PI-PLC that has an N-terminal pleckstrin homology (PH) domain, an array of EF hands, a PLC catalytic core domain, a C2 domain, and a unique C-terminal tail that terminates with a PDZ-binding motif, a potential interaction site for other signaling proteins. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. PI-PLC-eta2 is a neuron-specific enzyme and expressed in the brain. It may in part function downstream of G-protein-coupled receptors and play an important role in the formation and maintenance of the neuronal network in the postnatal brain. Pssm-ID: 176570 [Multi-domain] Cd Length: 254 Bit Score: 258.43 E-value: 1.93e-82
|
||||||||||||
| PI-PLCc_eta | cd08594 | Catalytic domain of metazoan phosphoinositide-specific phospholipase C-eta; This family ... |
52-379 | 6.97e-82 | ||||||||
Catalytic domain of metazoan phosphoinositide-specific phospholipase C-eta; This family corresponds to the catalytic domain present in metazoan phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11)-eta isozymes. PI-PLC is a signaling enzyme that hydrolyzes the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, Inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which then phosphorylates other molecules, leading to altered cellular activity. Calcium is required for the catalysis. PI-PLC-eta represents a class of neuron-speific PI-PLC that has an N-terminal pleckstrin homology (PH) domain, an array of EF hands, a PLC catalytic core domain, a C2 domain, and a unique C-terminal tail that terminates with a PDZ-binding motif, a potential interaction site for other signaling proteins. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. There are two PI-PLC-eta isozymes (1-2), both neuron-specific enzymes. They function as calcium sensors that are activated by small increases in intracellular calcium concentrations. The PI-PLC-eta isozymes are also activated through GPCR stimulation. Aside from the PI-PLC-eta isozymes identified in mammals, their eukaryotic homologs are also present in this family. Pssm-ID: 176536 [Multi-domain] Cd Length: 227 Bit Score: 255.88 E-value: 6.97e-82
|
||||||||||||
| PI-PLCc_epsilon | cd08596 | Catalytic domain of metazoan phosphoinositide-specific phospholipase C-epsilon; This family ... |
53-379 | 1.39e-81 | ||||||||
Catalytic domain of metazoan phosphoinositide-specific phospholipase C-epsilon; This family corresponds to the catalytic domain present in metazoan phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11)-epsilon isozymes. PI-PLC is a signaling enzyme that hydrolyzes the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which then phosphorylates other molecules, leading to altered cellular activity. Calcium is required for the catalysis. PI-PLC-epsilon represents a class of mammalian PI-PLC that has an N-terminal CDC25 homology domain with a guanyl-nucleotide exchange factor (GFF) activity, a pleckstrin homology (PH) domain, an array of EF hands, a PLC catalytic core domain, a C2 domain, and two predicted RA (Ras association) domains that are implicated in the binding of small GTPases, such as Ras or Rap, from the Ras family. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. There is one PI-PLC-epsilon isozyme (1). PI-PLC-epsilon is activated by G alpha(12/13), G beta gamma, and activated members of Ras and Rho small GTPases. Aside from PI-PLC-epsilon identified in mammals, its eukaryotic homologs have been classified with this family. Pssm-ID: 176538 [Multi-domain] Cd Length: 254 Bit Score: 256.31 E-value: 1.39e-81
|
||||||||||||
| PI-PLC-X | pfam00388 | Phosphatidylinositol-specific phospholipase C, X domain; This associates with pfam00387 to ... |
55-196 | 9.24e-81 | ||||||||
Phosphatidylinositol-specific phospholipase C, X domain; This associates with pfam00387 to form a single structural unit. Pssm-ID: 459795 [Multi-domain] Cd Length: 142 Bit Score: 249.73 E-value: 9.24e-81
|
||||||||||||
| PI-PLCc_eta1 | cd08632 | Catalytic domain of metazoan phosphoinositide-specific phospholipase C-eta1; This subfamily ... |
52-379 | 1.08e-80 | ||||||||
Catalytic domain of metazoan phosphoinositide-specific phospholipase C-eta1; This subfamily corresponds to the catalytic domain present in metazoan phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11)-eta isozyme 1. PI-PLC is a signaling enzyme that hydrolyzes the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, Inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which then phosphorylates other molecules, leading to altered cellular activity. Calcium is required for the catalysis. PI-PLC-eta represents a class of neuron-speific PI-PLC that has an N-terminal pleckstrin homology (PH) domain, an array of EF hands, a PLC catalytic core domain, a C2 domain, and a unique C-terminal tail that terminates with a PDZ-binding motif, a potential interaction site for other signaling proteins. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. PI-PLC-eta1 is a neuron-specific enzyme and expressed in only nerve tissues such as the brain and spinal cord. It may perform a fundamental role in the brain. Pssm-ID: 176569 [Multi-domain] Cd Length: 253 Bit Score: 253.80 E-value: 1.08e-80
|
||||||||||||
| PI-PLCc_beta4 | cd08626 | Catalytic domain of metazoan phosphoinositide-specific phospholipase C-beta4; This subfamily ... |
52-376 | 8.98e-79 | ||||||||
Catalytic domain of metazoan phosphoinositide-specific phospholipase C-beta4; This subfamily corresponds to the catalytic domain present in metazoan phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11)-beta isozyme 4. PI-PLC is a signaling enzyme that hydrolyzes the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, Inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which goes on to phosphorylate other molecules, leading to altered cellular activity. Calcium is required for the catalysis. PLC-beta represents a class of mammalian PI-PLC that has an N-terminal pleckstrin homology (PH) domain, an array of EF hands, a PLC catalytic core domain, a C2 domain, and a unique C-terminal coiled-coil (CT) domain necessary for homodimerization. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. PI-PLC-beta4 is expressed in high concentrations in cerebellar Purkinje and granule cells, the median geniculate body, and the lateral geniculate nucleus. It is activated by the heterotrimeric G protein alpha q subunits through their C2 domain and long C-terminal extension. Pssm-ID: 176563 [Multi-domain] Cd Length: 257 Bit Score: 248.91 E-value: 8.98e-79
|
||||||||||||
| PI-PLCc_gamma2 | cd08628 | Catalytic domain of metazoan phosphoinositide-specific phospholipase C-gamma2; This subfamily ... |
53-376 | 2.36e-77 | ||||||||
Catalytic domain of metazoan phosphoinositide-specific phospholipase C-gamma2; This subfamily corresponds to the catalytic domain present in metazoan phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11)-gamma isozyme 2. PI-PLC is a signaling enzyme that hydrolyze the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, Inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which goes on to phosphorylate other molecules, leading to altered cellular activity. Calcium is required for the catalysis. PI-PLC-gamma represents a class of mammalian PI-PLC that has an N-terminal pleckstrin homology (PH) domain, an array of EF hands, a PLC catalytic core domain, and a C2 domain. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. Unique to PI-PLC-gamma2, a second PH domain, two SH2 (Src homology 2) regions, and one SH3 (Src homology 3) region is present within this linker region. PI-PLC-gamma2 is highly expressed in cells of hematopoietic origin. It is activated by receptor and non-receptor tyrosine kinases due to the presence of two SH2 and a single SH3 domain within the linker region. Unlike PI-PLC-gamma1, the activation of PI-PLC-gamma2 may require concurrent stimulation of PI 3-kinase. Pssm-ID: 176565 [Multi-domain] Cd Length: 254 Bit Score: 245.35 E-value: 2.36e-77
|
||||||||||||
| PI-PLCc_beta2 | cd08624 | Catalytic domain of metazoan phosphoinositide-specific phospholipase C-beta2; This subfamily ... |
52-376 | 1.78e-73 | ||||||||
Catalytic domain of metazoan phosphoinositide-specific phospholipase C-beta2; This subfamily corresponds to the catalytic domain present in metazoan phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11)-beta isozyme 2. PI-PLC is a signaling enzyme that hydrolyzes the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, Inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which goes on to phosphorylate other molecules, leading to altered cellular activity. Calcium is required for the catalysis. PLC-beta represents a class of mammalian PI-PLC that has an N-terminal pleckstrin homology (PH) domain, an array of EF hands, a PLC catalytic core domain, a C2 domain, and a unique C-terminal coiled-coil (CT) domain necessary for homodimerization. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. PI-PLC-beta2 is expressed at highest levels in cells of hematopoietic origin. It is activated by the heterotrimeric G protein alpha q subunits through their C2 domain and long C-terminal extension. It is also activated by the beta-gamma subunits of heterotrimeric G proteins. Pssm-ID: 176561 [Multi-domain] Cd Length: 261 Bit Score: 235.34 E-value: 1.78e-73
|
||||||||||||
| PI-PLCc_beta3 | cd08625 | Catalytic domain of metazoan phosphoinositide-specific phospholipase C-beta3; This subfamily ... |
54-376 | 1.56e-71 | ||||||||
Catalytic domain of metazoan phosphoinositide-specific phospholipase C-beta3; This subfamily corresponds to the catalytic domain present in metazoan phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11)-beta isozyme 3. PI-PLC is a signaling enzyme that hydrolyzes the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, Inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which goes on to phosphorylate other molecules, leading to altered cellular activity. Calcium is required for the catalysis. PLC-beta represents a class of mammalian PI-PLC that has an N-terminal pleckstrin homology (PH) domain, an array of EF hands, a PLC catalytic core domain, a C2 domain, and a unique C-terminal coiled-coil (CT) domain necessary for homodimerization. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. PI-PLC-beta3 is widely expressed at highest levels in brain, liver, and parotid gland. It is activated by the heterotrimeric G protein alpha q subunits through their C2 domain and long C-terminal extension. It is also activated by the beta-gamma subunits of heterotrimeric G proteins. Pssm-ID: 176562 [Multi-domain] Cd Length: 258 Bit Score: 230.33 E-value: 1.56e-71
|
||||||||||||
| PI-PLCc_gamma1 | cd08627 | Catalytic domain of metazoan phosphoinositide-specific phospholipase C-gamma1; This subfamily ... |
53-379 | 5.82e-68 | ||||||||
Catalytic domain of metazoan phosphoinositide-specific phospholipase C-gamma1; This subfamily corresponds to the catalytic domain present in metazoan phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11)-gamma isozyme 1. PI-PLC is a signaling enzyme that hydrolyzes the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, Inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which goes on to phosphorylate other molecules, leading to altered cellular activity. Calcium is required for the catalysis. PI-PLC-gamma represents a class of mammalian PI-PLC that has an N-terminal pleckstrin homology (PH) domain, an array of EF hands, a PLC catalytic core domain, and a C2 domain. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. Unique to PI-PLC-gamma1, a second PH domain, two SH2 (Src homology 2) regions, and one SH3 (Src homology 3) region is present within this linker region. PI-PLC-gamma1 is ubiquitously expressed. It is activated by receptor and non-receptor tyrosine kinases due to the presence of two SH2 and a single SH3 domain within the linker region. Pssm-ID: 176564 [Multi-domain] Cd Length: 229 Bit Score: 219.90 E-value: 5.82e-68
|
||||||||||||
| PLN02952 | PLN02952 | phosphoinositide phospholipase C |
51-504 | 4.79e-67 | ||||||||
phosphoinositide phospholipase C Pssm-ID: 178538 [Multi-domain] Cd Length: 599 Bit Score: 228.73 E-value: 4.79e-67
|
||||||||||||
| PI-PLCc_beta1 | cd08623 | Catalytic domain of metazoan phosphoinositide-specific phospholipase C-beta1; This subfamily ... |
53-376 | 2.79e-64 | ||||||||
Catalytic domain of metazoan phosphoinositide-specific phospholipase C-beta1; This subfamily corresponds to the catalytic domain present in metazoan phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11)-beta isozyme 1. PI-PLC is a signaling enzyme that hydrolyzes the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, Inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which goes on to phosphorylate other molecules, leading to altered cellular activity. Calcium is required for the catalysis. PLC-beta represents a class of mammalian PI-PLC that has an N-terminal pleckstrin homology (PH) domain, an array of EF hands, a PLC catalytic core domain, a C2 domain, and a unique C-terminal coiled-coil (CT) domain necessary for homodimerization. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. PI-PLC-beta1 is expressed at highest levels in specific regions of the brain. It is activated by the heterotrimeric G protein alpha q subunits through their C2 domain and long C-terminal extension. Pssm-ID: 176560 [Multi-domain] Cd Length: 258 Bit Score: 211.48 E-value: 2.79e-64
|
||||||||||||
| PLN02228 | PLN02228 | Phosphoinositide phospholipase C |
29-503 | 7.96e-64 | ||||||||
Phosphoinositide phospholipase C Pssm-ID: 177873 [Multi-domain] Cd Length: 567 Bit Score: 219.14 E-value: 7.96e-64
|
||||||||||||
| PLCXc | smart00148 | Phospholipase C, catalytic domain (part); domain X; Phosphoinositide-specific phospholipases C. ... |
55-197 | 5.12e-61 | ||||||||
Phospholipase C, catalytic domain (part); domain X; Phosphoinositide-specific phospholipases C. These enzymes contain 2 regions (X and Y) which together form a TIM barrel-like structure containing the active site residues. Phospholipase C enzymes (PI-PLC) act as signal transducers that generate two second messengers, inositol-1,4,5-trisphosphate and diacylglycerol. The bacterial enzyme appears to be a homologue of the mammalian PLCs. Pssm-ID: 197543 [Multi-domain] Cd Length: 143 Bit Score: 198.27 E-value: 5.12e-61
|
||||||||||||
| PLN02222 | PLN02222 | phosphoinositide phospholipase C 2 |
7-503 | 1.19e-59 | ||||||||
phosphoinositide phospholipase C 2 Pssm-ID: 177868 [Multi-domain] Cd Length: 581 Bit Score: 208.35 E-value: 1.19e-59
|
||||||||||||
| PLN02230 | PLN02230 | phosphoinositide phospholipase C 4 |
25-503 | 3.01e-58 | ||||||||
phosphoinositide phospholipase C 4 Pssm-ID: 177875 [Multi-domain] Cd Length: 598 Bit Score: 204.94 E-value: 3.01e-58
|
||||||||||||
| PI-PLCc_plant | cd08599 | Catalytic domain of plant phosphatidylinositide-specific phospholipases C; This family ... |
52-379 | 1.27e-57 | ||||||||
Catalytic domain of plant phosphatidylinositide-specific phospholipases C; This family corresponds to the catalytic domain present in a group of phosphoinositide-specific phospholipases C (PI-PLC, EC 3.1.4.11) encoded by PLC genes from higher plants, which are homologs of mammalian PI-PLC in terms of overall sequence similarity and domain organization. Mammalian PI-PLC is a signaling enzyme that hydrolyzes the membrane phospholipids phosphatidylinositol-4,5-bisphosphate (PIP2) to generate two important second messengers in eukaryotic signal transduction cascades, inositol 1,4,5-trisphosphate (InsP3) and diacylglycerol (DAG). InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which then phosphorylates other molecules, leading to altered cellular activity. Calcium is required for the catalysis. The domain arrangement of plant PI-PLCs is structurally similar to the mammalian PLC-zeta isoform, which lacks the N-terminal pleckstrin homology (PH) domain, but contains EF-hand like motifs (which are absent in a few plant PLCs), a PLC catalytic core domain with X- and Y- highly conserved regions split by a linker sequence, and a C2 domain. However, at the sequence level, the plant PI-PLCs are closely related to the mammalian PLC-delta isoform. Experiments show that plant PLCs display calcium dependent PLC catalytic properties, although they lack some of the N-terminal motifs found in their mammalian counterparts. A putative calcium binding site may be located at the region spanning the X- and Y- domains. Pssm-ID: 176541 [Multi-domain] Cd Length: 228 Bit Score: 192.59 E-value: 1.27e-57
|
||||||||||||
| PI-PLC-Y | pfam00387 | Phosphatidylinositol-specific phospholipase C, Y domain; This associates with pfam00388 to ... |
276-389 | 5.03e-57 | ||||||||
Phosphatidylinositol-specific phospholipase C, Y domain; This associates with pfam00388 to form a single structural unit. Pssm-ID: 459794 Cd Length: 114 Bit Score: 186.90 E-value: 5.03e-57
|
||||||||||||
| PI-PLCc | cd00137 | Catalytic domain of prokaryotic and eukaryotic phosphoinositide-specific phospholipase C; This ... |
52-379 | 7.23e-54 | ||||||||
Catalytic domain of prokaryotic and eukaryotic phosphoinositide-specific phospholipase C; This subfamily corresponds to the catalytic domain present in prokaryotic and eukaryotic phosphoinositide-specific phospholipase C (PI-PLC), which is a ubiquitous enzyme catalyzing the cleavage of the sn3-phosphodiester bond in the membrane phosphoinositides (phosphatidylinositol, PI; Phosphatidylinositol-4-phosphate, PIP; phosphatidylinositol 4,5-bisphosphate, PIP2) to yield inositol phosphates (inositol monosphosphate, InsP; inositol diphosphate, InsP2; inositol trisphosphate, InsP3) and diacylglycerol (DAG). The higher eukaryotic PI-PLCs (EC 3.1.4.11) have a multidomain organization that consists of a PLC catalytic core domain, and various regulatory domains. They play a critical role in most signal transduction pathways, controlling numerous cellular events, such as cell growth, proliferation, excitation and secretion. These PI-PLCs strictly require Ca2+ for their catalytic activity. They display a clear preference towards the hydrolysis of the more highly phosphorylated PI-analogues, PIP2 and PIP, to generate two important second messengers, InsP3 and DAG. InsP3 triggers inflow of calcium from intracellular stores, while DAG, together with calcium, activates protein kinase C, which then phosphorylates other molecules, leading to altered cellular activity. In contrast, bacterial PI-PLCs contain a single catalytic domain. Although their precise physiological function remains unclear, bacterial PI-PLCs may function as virulence factors in some pathogenic bacteria. They participate in Ca2+-independent PI metabolism. They are characterized as phosphatidylinositol-specific phospholipase C (EC 4.6.1.13) that selectively hydrolyze PI, not PIP or PIP2. The TIM-barrel type catalytic domain in bacterial PI-PLCs is very similar to the one in eukaryotic PI-PLCs, in which the catalytic domain is assembled from two highly conserved X- and Y-regions split by a divergent linker sequence. The catalytic mechanism of both prokaryotic and eukaryotic PI-PLCs is based on general base and acid catalysis utilizing two well conserved histidines, and consists of two steps, a phosphotransfer and a phosphodiesterase reaction. This superfamily also includes a distinctly different type of eukaryotic PLC, glycosylphosphatidylinositol-specific phospholipase C (GPI-PLC), an integral membrane protein characterized in the protozoan parasite Trypanosoma brucei. T. brucei GPI-PLC hydrolyzes the GPI-anchor on the variant specific glycoprotein (VSG), releasing dimyristyl glycerol (DMG), which may facilitate the evasion of the protozoan to the host#s immune system. It does not require Ca2+ for its activity and is more closely related to bacterial PI-PLCs, but not mammalian PI-PLCs. Pssm-ID: 176497 [Multi-domain] Cd Length: 274 Bit Score: 184.39 E-value: 7.23e-54
|
||||||||||||
| PLCYc | smart00149 | Phospholipase C, catalytic domain (part); domain Y; Phosphoinositide-specific phospholipases C. ... |
277-391 | 1.11e-46 | ||||||||
Phospholipase C, catalytic domain (part); domain Y; Phosphoinositide-specific phospholipases C. These enzymes contain 2 regions (X and Y) which together form a TIM barrel-like structure containing the active site residues. Phospholipase C enzymes (PI-PLC) act as signal transducers that generate two second messengers, inositol-1,4,5-trisphosphate and diacylglycerol. The bacterial enzyme appears to be a homologue of the mammalian PLCs. Pssm-ID: 128454 [Multi-domain] Cd Length: 115 Bit Score: 159.33 E-value: 1.11e-46
|
||||||||||||
| PLN02223 | PLN02223 | phosphoinositide phospholipase C |
52-503 | 3.69e-39 | ||||||||
phosphoinositide phospholipase C Pssm-ID: 165867 [Multi-domain] Cd Length: 537 Bit Score: 150.56 E-value: 3.69e-39
|
||||||||||||
| C2_PLC_like | cd00275 | C2 domain present in Phosphoinositide-specific phospholipases C (PLC); PLCs are involved in ... |
409-527 | 3.73e-32 | ||||||||
C2 domain present in Phosphoinositide-specific phospholipases C (PLC); PLCs are involved in the hydrolysis of phosphatidylinositol-4,5-bisphosphate (PIP2) to d-myo-inositol-1,4,5-trisphosphate (1,4,5-IP3) and sn-1,2-diacylglycerol (DAG). 1,4,5-IP3 and DAG are second messengers in eukaryotic signal transduction cascades. PLC is composed of a N-terminal PH domain followed by a series of EF hands, a catalytic TIM barrel and a C-terminal C2 domain. C2 domains fold into an 8-standed beta-sandwich that can adopt 2 structural arrangements: Type I and Type II, distinguished by a circular permutation involving their N- and C-terminal beta strands. Many C2 domains are Ca2+-dependent membrane-targeting modules that bind a wide variety of substances including bind phospholipids, inositol polyphosphates, and intracellular proteins. Most C2 domain proteins are either signal transduction enzymes that contain a single C2 domain, such as protein kinase C, or membrane trafficking proteins which contain at least two C2 domains, such as synaptotagmin 1. However, there are a few exceptions to this including RIM isoforms and some splice variants of piccolo/aczonin and intersectin which only have a single C2 domain. C2 domains with a calcium binding region have negatively charged residues, primarily aspartates, that serve as ligands for calcium ions. Members here have a type-II topology. Pssm-ID: 175974 [Multi-domain] Cd Length: 128 Bit Score: 120.34 E-value: 3.73e-32
|
||||||||||||
| C2 | smart00239 | Protein kinase C conserved region 2 (CalB); Ca2+-binding motif present in phospholipases, ... |
411-514 | 5.79e-18 | ||||||||
Protein kinase C conserved region 2 (CalB); Ca2+-binding motif present in phospholipases, protein kinases C, and synaptotagmins (among others). Some do not appear to contain Ca2+-binding sites. Particular C2s appear to bind phospholipids, inositol polyphosphates, and intracellular proteins. Unusual occurrence in perforin. Synaptotagmin and PLC C2s are permuted in sequence with respect to N- and C-terminal beta strands. SMART detects C2 domains using one or both of two profiles. Pssm-ID: 214577 [Multi-domain] Cd Length: 101 Bit Score: 79.45 E-value: 5.79e-18
|
||||||||||||
| PI-PLCc_GDPD_SF | cd08555 | Catalytic domain of phosphoinositide-specific phospholipase C-like phosphodiesterases ... |
66-171 | 9.93e-16 | ||||||||
Catalytic domain of phosphoinositide-specific phospholipase C-like phosphodiesterases superfamily; The PI-PLC-like phosphodiesterases superfamily represents the catalytic domains of bacterial phosphatidylinositol-specific phospholipase C (PI-PLC, EC 4.6.1.13), eukaryotic phosphoinositide-specific phospholipase C (PI-PLC, EC 3.1.4.11), glycerophosphodiester phosphodiesterases (GP-GDE, EC 3.1.4.46), sphingomyelinases D (SMases D) (sphingomyelin phosphodiesterase D, EC 3.1.4.41) from spider venom, SMases D-like proteins, and phospholipase D (PLD) from several pathogenic bacteria, as well as their uncharacterized homologs found in organisms ranging from bacteria and archaea to metazoans, plants, and fungi. PI-PLCs are ubiquitous enzymes hydrolyzing the membrane lipid phosphoinositides to yield two important second messengers, inositol phosphates and diacylglycerol (DAG). GP-GDEs play essential roles in glycerol metabolism and catalyze the hydrolysis of glycerophosphodiesters to sn-glycerol-3-phosphate (G3P) and the corresponding alcohols that are major sources of carbon and phosphate. Both, PI-PLCs and GP-GDEs, can hydrolyze the 3'-5' phosphodiester bonds in different substrates, and utilize a similar mechanism of general base and acid catalysis with conserved histidine residues, which consists of two steps, a phosphotransfer and a phosphodiesterase reaction. This superfamily also includes Neurospora crassa ankyrin repeat protein NUC-2 and its Saccharomyces cerevisiae counterpart, Phosphate system positive regulatory protein PHO81, glycerophosphodiester phosphodiesterase (GP-GDE)-like protein SHV3 and SHV3-like proteins (SVLs). The residues essential for enzyme activities and metal binding are not conserved in these sequence homologs, which might suggest that the function of catalytic domains in these proteins might be distinct from those in typical PLC-like phosphodiesterases. Pssm-ID: 176498 [Multi-domain] Cd Length: 179 Bit Score: 75.55 E-value: 9.93e-16
|
||||||||||||
| C2 | pfam00168 | C2 domain; |
411-503 | 1.56e-15 | ||||||||
C2 domain; Pssm-ID: 425499 [Multi-domain] Cd Length: 104 Bit Score: 72.35 E-value: 1.56e-15
|
||||||||||||
| C2 | cd00030 | C2 domain; The C2 domain was first identified in PKC. C2 domains fold into an 8-standed ... |
412-503 | 8.89e-13 | ||||||||
C2 domain; The C2 domain was first identified in PKC. C2 domains fold into an 8-standed beta-sandwich that can adopt 2 structural arrangements: Type I and Type II, distinguished by a circular permutation involving their N- and C-terminal beta strands. Many C2 domains are Ca2+-dependent membrane-targeting modules that bind a wide variety of substances including bind phospholipids, inositol polyphosphates, and intracellular proteins. Most C2 domain proteins are either signal transduction enzymes that contain a single C2 domain, such as protein kinase C, or membrane trafficking proteins which contain at least two C2 domains, such as synaptotagmin 1. However, there are a few exceptions to this including RIM isoforms and some splice variants of piccolo/aczonin and intersectin which only have a single C2 domain. C2 domains with a calcium binding region have negatively charged residues, primarily aspartates, that serve as ligands for calcium ions. Pssm-ID: 175973 [Multi-domain] Cd Length: 102 Bit Score: 64.40 E-value: 8.89e-13
|
||||||||||||
| EFh_PI-PLCzeta | cd16204 | EF-hand motif found in phosphoinositide phospholipase C zeta 1 (PI-PLC-zeta1); PI-PLC-zeta1, ... |
2-40 | 2.36e-08 | ||||||||
EF-hand motif found in phosphoinositide phospholipase C zeta 1 (PI-PLC-zeta1); PI-PLC-zeta1, also termed 1-phosphatidylinositol 4,5-bisphosphate phosphodiesterase zeta-1, or phospholipase C-zeta-1 (PLC-zeta-1), or testis-development protein NYD-SP27, is only found in the testis. The sperm-specific PI-PLC plays a fundamental role in vertebrate fertilization by initiating intracellular calcium oscillations that trigger the embryo development. However, the mechanism of its activation still remains unclear. PI-PLC-zeta1 contains an N-terminal four atypical EF-hand motifs, a PLC catalytic core domain, and a C-terminal C2 domain. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. Unlike other PI-PLCs, PI-PLC-zeta is responsible for Ca2+ oscillations in fertilized oocytes and exhibits a high sensitivity to Ca2+ mediated through its EF-hand domain. There is only one PLC-zeta isozyme. Aside from PI-PLC-zeta identified in mammals, its eukaryotic homologs have been classified with this family. Pssm-ID: 320034 [Multi-domain] Cd Length: 142 Bit Score: 53.28 E-value: 2.36e-08
|
||||||||||||
| C2B_Synaptotagmin | cd00276 | C2 domain second repeat present in Synaptotagmin; Synaptotagmin is a membrane-trafficking ... |
408-498 | 2.49e-07 | ||||||||
C2 domain second repeat present in Synaptotagmin; Synaptotagmin is a membrane-trafficking protein characterized by a N-terminal transmembrane region, a linker, and 2 C-terminal C2 domains. There are several classes of Synaptotagmins. Previously all synaptotagmins were thought to be calcium sensors in the regulation of neurotransmitter release and hormone secretion, but it has been shown that not all of them bind calcium. Of the 17 identified synaptotagmins only 8 bind calcium (1-3, 5-7, 9, 10). The function of the two C2 domains that bind calcium are: regulating the fusion step of synaptic vesicle exocytosis (C2A) and binding to phosphatidyl-inositol-3,4,5-triphosphate (PIP3) in the absence of calcium ions and to phosphatidylinositol bisphosphate (PIP2) in their presence (C2B). C2B also regulates also the recycling step of synaptic vesicles. C2 domains fold into an 8-standed beta-sandwich that can adopt 2 structural arrangements: Type I and Type II, distinguished by a circular permutation involving their N- and C-terminal beta strands. Many C2 domains are Ca2+-dependent membrane-targeting modules that bind a wide variety of substances including bind phospholipids, inositol polyphosphates, and intracellular proteins. Most C2 domain proteins are either signal transduction enzymes that contain a single C2 domain, such as protein kinase C, or membrane trafficking proteins which contain at least two C2 domains, such as synaptotagmin 1. However, there are a few exceptions to this including RIM isoforms and some splice variants of piccolo/aczonin and intersectin which only have a single C2 domain. C2 domains with a calcium binding region have negatively charged residues, primarily aspartates, that serve as ligands for calcium ions. This cd contains the second C2 repeat, C2B, and has a type-I topology. Pssm-ID: 175975 [Multi-domain] Cd Length: 134 Bit Score: 49.89 E-value: 2.49e-07
|
||||||||||||
| C2B_Synaptotagmin-like | cd04050 | C2 domain second repeat present in Synaptotagmin-like proteins; Synaptotagmin is a ... |
412-503 | 7.50e-07 | ||||||||
C2 domain second repeat present in Synaptotagmin-like proteins; Synaptotagmin is a membrane-trafficking protein characterized by a N-terminal transmembrane region, a linker, and 2 C-terminal C2 domains. Previously all synaptotagmins were thought to be calcium sensors in the regulation of neurotransmitter release and hormone secretion, but it has been shown that not all of them bind calcium. Of the 17 identified synaptotagmins only 8 bind calcium (1-3, 5-7, 9, 10). The function of the two C2 domains that bind calcium are: regulating the fusion step of synaptic vesicle exocytosis (C2A) and binding to phosphatidyl-inositol-3,4,5-triphosphate (PIP3) in the absence of calcium ions and to phosphatidylinositol bisphosphate (PIP2) in their presence (C2B). C2B also regulates also the recycling step of synaptic vesicles. C2 domains fold into an 8-standed beta-sandwich that can adopt 2 structural arrangements: Type I and Type II, distinguished by a circular permutation involving their N- and C-terminal beta strands. Many C2 domains are Ca2+-dependent membrane-targeting modules that bind a wide variety of substances including bind phospholipids, inositol polyphosphates, and intracellular proteins. Most C2 domain proteins are either signal transduction enzymes that contain a single C2 domain, such as protein kinase C, or membrane trafficking proteins which contain at least two C2 domains, such as synaptotagmin 1. However, there are a few exceptions to this including RIM isoforms and some splice variants of piccolo/aczonin and intersectin which only have a single C2 domain. C2 domains with a calcium binding region have negatively charged residues, primarily aspartates, that serve as ligands for calcium ions. This cd contains the second C2 repeat, C2B, and has a type-I topology. Pssm-ID: 176015 [Multi-domain] Cd Length: 105 Bit Score: 47.94 E-value: 7.50e-07
|
||||||||||||
| PI-PLCc_bacteria_like | cd08557 | Catalytic domain of bacterial phosphatidylinositol-specific phospholipase C and similar ... |
54-171 | 2.60e-06 | ||||||||
Catalytic domain of bacterial phosphatidylinositol-specific phospholipase C and similar proteins; This subfamily corresponds to the catalytic domain present in bacterial phosphatidylinositol-specific phospholipase C (PI-PLC, EC 4.6.1.13) and their sequence homologs found in eukaryota. Bacterial PI-PLCs participate in Ca2+-independent PI metabolism, hydrolyzing the membrane lipid phosphatidylinositol (PI) to produce phosphorylated myo-inositol and diacylglycerol (DAG). Although their precise physiological function remains unclear, bacterial PI-PLCs may function as virulence factors in some pathogenic bacteria. Bacterial PI-PLCs contain a single TIM-barrel type catalytic domain. Its catalytic mechanism is based on general base and acid catalysis utilizing two well conserved histidines, and consists of two steps, a phosphotransfer and a phosphodiesterase reaction. Eukaryotic homologs in this family are named as phosphatidylinositol-specific phospholipase C X domain containing proteins (PI-PLCXD). They are distinct from the typical eukaryotic phosphoinositide-specific phospholipases C (PI-PLC, EC 3.1.4.11), which have a multidomain organization that consists of a PLC catalytic core domain, and various regulatory domains. The catalytic core domain is assembled from two highly conserved X- and Y-regions split by a divergent linker sequence. In contrast, eukaryotic PI-PLCXDs contain a single TIM-barrel type catalytic domain, X domain, which is closely related to that of bacterial PI-PLCs. Although the biological function of eukaryotic PI-PLCXDs still remains unclear, it may be distinct from that of typical eukaryotic PI-PLCs. This family also includes a distinctly different type of eukaryotic PLC, glycosylphosphatidylinositol-specific phospholipase C (GPI-PLC), an integral membrane protein characterized in the protozoan parasite Trypanosoma brucei. T. brucei GPI-PLC hydrolyzes the GPI-anchor on the variant specific glycoprotein (VSG), releasing dimyristyl glycerol (DMG), which may facilitate the evasion of the protozoan to the host's immune system. It does not require Ca2+ for its activity and is more closely related to bacterial PI-PLCs, but not mammalian PI-PLCs. Pssm-ID: 176500 [Multi-domain] Cd Length: 271 Bit Score: 49.40 E-value: 2.60e-06
|
||||||||||||
| C2C_KIAA1228 | cd04030 | C2 domain third repeat present in uncharacterized human KIAA1228-like proteins; KIAA proteins ... |
421-498 | 3.20e-06 | ||||||||
C2 domain third repeat present in uncharacterized human KIAA1228-like proteins; KIAA proteins are uncharacterized human proteins. They were compiled by the Kazusa mammalian cDNA project which identified more than 2000 human genes. They are identified by 4 digit codes that precede the KIAA designation. Many KIAA genes are still functionally uncharacterized including KIAA1228. C2 domains fold into an 8-standed beta-sandwich that can adopt 2 structural arrangements: Type I and Type II, distinguished by a circular permutation involving their N- and C-terminal beta strands. Many C2 domains are Ca2+-dependent membrane-targeting modules that bind a wide variety of substances including bind phospholipids, inositol polyphosphates, and intracellular proteins. Most C2 domain proteins are either signal transduction enzymes that contain a single C2 domain, such as protein kinase C, or membrane trafficking proteins which contain at least two C2 domains, such as synaptotagmin 1. However, there are a few exceptions to this including RIM isoforms and some splice variants of piccolo/aczonin and intersectin which only have a single C2 domain. C2 domains with a calcium binding region have negatively charged residues, primarily aspartates, that serve as ligands for calcium ions. This cd contains the third C2 repeat, C2C, and has a type-II topology. Pssm-ID: 175996 [Multi-domain] Cd Length: 127 Bit Score: 46.50 E-value: 3.20e-06
|
||||||||||||
| C2_cPLA2 | cd04036 | C2 domain present in cytosolic PhosphoLipase A2 (cPLA2); A single copy of the C2 domain is ... |
412-472 | 5.35e-06 | ||||||||
C2 domain present in cytosolic PhosphoLipase A2 (cPLA2); A single copy of the C2 domain is present in cPLA2 which releases arachidonic acid from membranes initiating the biosynthesis of potent inflammatory mediators such as prostaglandins, leukotrienes, and platelet-activating factor. C2 domains fold into an 8-standed beta-sandwich that can adopt 2 structural arrangements: Type I and Type II, distinguished by a circular permutation involving their N- and C-terminal beta strands. Many C2 domains are Ca2+-dependent membrane-targeting modules that bind a wide variety of substances including bind phospholipids, inositol polyphosphates, and intracellular proteins. Most C2 domain proteins are either signal transduction enzymes that contain a single C2 domain, such as protein kinase C, or membrane trafficking proteins which contain at least two C2 domains, such as synaptotagmin 1. However, there are a few exceptions to this including RIM isoforms and some splice variants of piccolo/aczonin and intersectin which only have a single C2 domain. C2 domains with a calcium binding region have negatively charged residues, primarily aspartates, that serve as ligands for calcium ions. Members of this cd have a type-II topology. Pssm-ID: 176001 [Multi-domain] Cd Length: 119 Bit Score: 45.72 E-value: 5.35e-06
|
||||||||||||
| EFh_PRIP | cd16206 | EF-hand motif found in phospholipase C-related but catalytically inactive proteins (PRIP); ... |
7-40 | 7.24e-06 | ||||||||
EF-hand motif found in phospholipase C-related but catalytically inactive proteins (PRIP); This family represents a class of metazoan phospholipase C related, but catalytically inactive proteins (PRIP), which belong to a group of novel inositol 1,4,5-trisphosphate (InsP3) binding protein. PRIP has a primary structure and domain architecture, incorporating a pleckstrin homology (PH) domain, four atypical EF-hand motifs, a PLC catalytic core domain with highly conserved X- and Y-regions split by a linker sequence, and a C-terminal C2 domain, similar to phosphoinositide-specific phospholipases C (PI-PLC, EC 3.1.4.11)-delta isoforms. Due to replacement of critical catalytic residues, PRIP do not have PLC enzymatic activity. PRIP consists of two subfamilies, PRIP-1(also known as p130 or PLC-L1), which is predominantly expressed in the brain, and PRIP-2 (also known as PLC-L2), which exhibits a relatively ubiquitous expression. Experiments show both, PRIP-1 and PRIP-2, are involved in InsP3-mediated calcium signaling pathway and GABA(A)receptor-mediated signaling pathway. In addition, PRIP-2 acts as a negative regulator of B-cell receptor signaling and immune responses. Pssm-ID: 320036 [Multi-domain] Cd Length: 143 Bit Score: 46.05 E-value: 7.24e-06
|
||||||||||||
| EF-hand_like | pfam09279 | Phosphoinositide-specific phospholipase C, efhand-like; Members of this family are ... |
1-44 | 1.72e-05 | ||||||||
Phosphoinositide-specific phospholipase C, efhand-like; Members of this family are predominantly found in phosphoinositide-specific phospholipase C. They adopt a structure consisting of a core of four alpha helices, in an EF like fold, and are required for functioning of the enzyme. Pssm-ID: 401279 [Multi-domain] Cd Length: 85 Bit Score: 43.39 E-value: 1.72e-05
|
||||||||||||
| C2A_Synaptotagmin-7 | cd08386 | C2A domain first repeat present in Synaptotagmin 7; Synaptotagmin is a membrane-trafficking ... |
404-469 | 2.64e-05 | ||||||||
C2A domain first repeat present in Synaptotagmin 7; Synaptotagmin is a membrane-trafficking protein characterized by a N-terminal transmembrane region, a linker, and 2 C-terminal C2 domains. Synaptotagmin 7, a member of class 2 synaptotagmins, is located in presynaptic plasma membranes in neurons, dense-core vesicles in endocrine cells, and lysosomes in fibroblasts. It has been shown to play a role in regulation of Ca2+-dependent lysosomal exocytosis in fibroblasts and may also function as a vesicular Ca2+-sensor. It is distinguished from the other synaptotagmins by having over 12 splice forms. Previously all synaptotagmins were thought to be calcium sensors in the regulation of neurotransmitter release and hormone secretion, but it has been shown that not all of them bind calcium. Of the 17 identified synaptotagmins only 8 bind calcium (1-3, 5-7, 9, 10). The function of the two C2 domains that bind calcium are: regulating the fusion step of synaptic vesicle exocytosis (C2A) and binding to phosphatidyl-inositol-3,4,5-triphosphate (PIP3) in the absence of calcium ions and to phosphatidylinositol bisphosphate (PIP2) in their presence (C2B). C2B also regulates also the recycling step of synaptic vesicles. C2 domains fold into an 8-standed beta-sandwich that can adopt 2 structural arrangements: Type I and Type II, distinguished by a circular permutation involving their N- and C-terminal beta strands. Many C2 domains are Ca2+-dependent membrane-targeting modules that bind a wide variety of substances including bind phospholipids, inositol polyphosphates, and intracellular proteins. Most C2 domain proteins are either signal transduction enzymes that contain a single C2 domain, such as protein kinase C, or membrane trafficking proteins which contain at least two C2 domains, such as synaptotagmin 1. However, there are a few exceptions to this including RIM isoforms and some splice variants of piccolo/aczonin and intersectin which only have a single C2 domain. C2 domains with a calcium binding region have negatively charged residues, primarily aspartates, that serve as ligands for calcium ions. This cd contains the first C2 repeat, C2A, and has a type-I topology. Pssm-ID: 176032 [Multi-domain] Cd Length: 125 Bit Score: 43.86 E-value: 2.64e-05
|
||||||||||||
| C2_Freud-1 | cd08690 | C2 domain found in 5' repressor element under dual repression binding protein-1 (Freud-1); ... |
412-498 | 4.23e-05 | ||||||||
C2 domain found in 5' repressor element under dual repression binding protein-1 (Freud-1); Freud-1 is a novel calcium-regulated repressor that negatively regulates basal 5-HT1A receptor expression in neurons. It may also play a role in the altered regulation of 5-HT1A receptors associated with anxiety or major depression. Freud-1 contains two DM-14 basic repeats, a helix-loop-helix DNA binding domain, and a C2 domain. The Freud-1 C2 domain is thought to be calcium insensitive and it lacks several acidic residues that mediate calcium binding of the PKC C2 domain. In addition, it contains a poly-basic insert that is not present in calcium-dependent C2 domains and may function as a nuclear localization signal. C2 domains fold into an 8-standed beta-sandwich that can adopt 2 structural arrangements: Type I and Type II, distinguished by a circular permutation involving their N- and C-terminal beta strands. Many C2 domains are Ca2+-dependent membrane-targeting modules that bind a wide variety of substances including bind phospholipids, inositol polyphosphates, and intracellular proteins. Most C2 domain proteins are either signal transduction enzymes that contain a single C2 domain, such as protein kinase C, or membrane trafficking proteins which contain at least two C2 domains, such as synaptotagmin 1. However, there are a few exceptions to this including RIM isoforms and some splice variants of piccolo/aczonin and intersectin which only have a single C2 domain. C2 domains with a calcium binding region have negatively charged residues, primarily aspartates, that serve as ligands for calcium ions. This cd contains the first C2 repeat, C2A, and has a type-II topology. Pssm-ID: 176072 [Multi-domain] Cd Length: 155 Bit Score: 43.84 E-value: 4.23e-05
|
||||||||||||
| C2A_Synaptotagmin-1-5-6-9-10 | cd08385 | C2A domain first repeat present in Synaptotagmins 1, 5, 6, 9, and 10; Synaptotagmin is a ... |
404-474 | 6.51e-05 | ||||||||
C2A domain first repeat present in Synaptotagmins 1, 5, 6, 9, and 10; Synaptotagmin is a membrane-trafficking protein characterized by a N-terminal transmembrane region, a linker, and 2 C-terminal C2 domains. Synaptotagmin 1, a member of class 1 synaptotagmins, is located in the brain and endocranium and localized to the synaptic vesicles and secretory granules. It functions as a Ca2+ sensor for fast exocytosis as do synaptotagmins 5, 6, and 10. It is distinguished from the other synaptotagmins by having an N-glycosylated N-terminus. Synaptotagmins 5, 6, and 10, members of class 3 synaptotagmins, are located primarily in the brain and localized to the active zone and plasma membrane. They is distinguished from the other synaptotagmins by having disulfide bonds at its N-terminus. Synaptotagmin 6 also regulates the acrosome reaction, a unique Ca2+-regulated exocytosis, in sperm. Synaptotagmin 9, a class 5 synaptotagmins, is located in the brain and localized to the synaptic vesicles. It is thought to be a Ca2+-sensor for dense-core vesicle exocytosis. Previously all synaptotagmins were thought to be calcium sensors in the regulation of neurotransmitter release and hormone secretion, but it has been shown that not all of them bind calcium. Of the 17 identified synaptotagmins only 8 bind calcium (1-3, 5-7, 9, 10). The function of the two C2 domains that bind calcium are: regulating the fusion step of synaptic vesicle exocytosis (C2A) and binding to phosphatidyl-inositol-3,4,5-triphosphate (PIP3) in the absence of calcium ions and to phosphatidylinositol bisphosphate (PIP2) in their presence (C2B). C2B also regulates also the recycling step of synaptic vesicles. C2 domains fold into an 8-standed beta-sandwich that can adopt 2 structural arrangements: Type I and Type II, distinguished by a circular permutation involving their N- and C-terminal beta strands. Many C2 domains are Ca2+-dependent membrane-targeting modules that bind a wide variety of substances including bind phospholipids, inositol polyphosphates, and intracellular proteins. Most C2 domain proteins are either signal transduction enzymes that contain a single C2 domain, such as protein kinase C, or membrane trafficking proteins which contain at least two C2 domains, such as synaptotagmin 1. However, there are a few exceptions to this including RIM isoforms and some splice variants of piccolo/aczonin and intersectin which only have a single C2 domain. C2 domains with a calcium binding region have negatively charged residues, primarily aspartates, that serve as ligands for calcium ions. This cd contains the first C2 repeat, C2A, and has a type-I topology. Pssm-ID: 176031 [Multi-domain] Cd Length: 124 Bit Score: 42.64 E-value: 6.51e-05
|
||||||||||||
| EFh_PI-PLCdelta | cd16202 | EF-hand motif found in phosphoinositide phospholipase C delta (PI-PLC-delta); PI-PLC-delta ... |
10-40 | 7.89e-05 | ||||||||
EF-hand motif found in phosphoinositide phospholipase C delta (PI-PLC-delta); PI-PLC-delta isozymes represent a class of metazoan PI-PLCs that are some of the most sensitive to calcium among all PLCs. Their activation is modulated by intracellular calcium ion concentration, phospholipids, polyamines, and other proteins, such as RhoAGAP. Like other PI-PLC isozymes, PI-PLC-delta isozymes contain a core set of domains, including an N-terminal pleckstrin homology (PH) domain, four atypical EF-hand motifs, a PLC catalytic core, and a single C-terminal C2 domain. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. There are three PI-PLC-delta isozymes (1, 3 and 4). PI-PLC-delta1 is relatively well characterized. It is activated by high calcium levels generated by other PI-PLC family members, and therefore functions as a calcium amplifier within the cell. Different PI-PLC-delta isozymes have different tissue distribution and different subcellular locations. PI-PLC-delta1 is mostly a cytoplasmic protein, PI-PLC-delta3 is located in the membrane, and PI-PLC-delta4 is predominantly detected in the cell nucleus. PI-PLC-delta isozymes is evolutionarily conserved even in non-mammalian species, such as yeast, slime molds and plants. Pssm-ID: 320032 [Multi-domain] Cd Length: 140 Bit Score: 42.98 E-value: 7.89e-05
|
||||||||||||
| C2A_RIM1alpha | cd04031 | C2 domain first repeat contained in Rab3-interacting molecule (RIM) proteins; RIMs are ... |
405-498 | 7.99e-05 | ||||||||
C2 domain first repeat contained in Rab3-interacting molecule (RIM) proteins; RIMs are believed to organize specialized sites of the plasma membrane called active zones. They also play a role in controlling neurotransmitter release, plasticity processes, as well as memory and learning. RIM contains an N-terminal zinc finger domain, a PDZ domain, and two C-terminal C2 domains (C2A, C2B). C2 domains fold into an 8-standed beta-sandwich that can adopt 2 structural arrangements: Type I and Type II, distinguished by a circular permutation involving their N- and C-terminal beta strands. Many C2 domains are Ca2+-dependent membrane-targeting modules that bind a wide variety of substances including bind phospholipids, inositol polyphosphates, and intracellular proteins. Most C2 domain proteins are either signal transduction enzymes that contain a single C2 domain, such as protein kinase C, or membrane trafficking proteins which contain at least two C2 domains, such as synaptotagmin 1. However, there are a few exceptions to this including RIM isoforms and some splice variants of piccolo/aczonin and intersectin which only have a single C2 domain. C2 domains with a calcium binding region have negatively charged residues, primarily aspartates, that serve as ligands for calcium ions. Members here have a type-I topology and do not bind Ca2+. Pssm-ID: 175997 [Multi-domain] Cd Length: 125 Bit Score: 42.62 E-value: 7.99e-05
|
||||||||||||
| C2A_Synaptotagmin-15-17 | cd08390 | C2A domain first repeat present in Synaptotagmins 15 and 17; Synaptotagmin is a ... |
404-507 | 1.87e-04 | ||||||||
C2A domain first repeat present in Synaptotagmins 15 and 17; Synaptotagmin is a membrane-trafficking protein characterized by a N-terminal transmembrane region, a linker, and 2 C-terminal C2 domains. It is thought to be involved in the trafficking and exocytosis of secretory vesicles in non-neuronal tissues and is Ca2+ independent. Human synaptotagmin 15 has 2 alternatively spliced forms that encode proteins with different C-termini. The larger, SYT15a, contains a N-terminal TM region, a putative fatty-acylation site, and 2 tandem C terminal C2 domains. The smaller, SYT15b, lacks the C-terminal portion of the second C2 domain. Unlike most other synaptotagmins it is nearly absent in the brain and rather is found in the heart, lungs, skeletal muscle, and testis. Synaptotagmin 17 is located in the brain, kidney, and prostate and is thought to be a peripheral membrane protein. Previously all synaptotagmins were thought to be calcium sensors in the regulation of neurotransmitter release and hormone secretion, but it has been shown that not all of them bind calcium. Of the 17 identified synaptotagmins only 8 bind calcium (1-3, 5-7, 9, 10). The function of the two C2 domains that bind calcium are: regulating the fusion step of synaptic vesicle exocytosis (C2A) and binding to phosphatidyl-inositol-3,4,5-triphosphate (PIP3) in the absence of calcium ions and to phosphatidylinositol bisphosphate (PIP2) in their presence (C2B). C2B also regulates also the recycling step of synaptic vesicles. C2 domains fold into an 8-standed beta-sandwich that can adopt 2 structural arrangements: Type I and Type II, distinguished by a circular permutation involving their N- and C-terminal beta strands. Many C2 domains are Ca2+-dependent membrane-targeting modules that bind a wide variety of substances including bind phospholipids, inositol polyphosphates, and intracellular proteins. Most C2 domain proteins are either signal transduction enzymes that contain a single C2 domain, such as protein kinase C, or membrane trafficking proteins which contain at least two C2 domains, such as synaptotagmin 1. However, there are a few exceptions to this including RIM isoforms and some splice variants of piccolo/aczonin and intersectin which only have a single C2 domain. C2 domains with a calcium binding region have negatively charged residues, primarily aspartates, that serve as ligands for calcium ions. This cd contains the first C2 repeat, C2A, and has a type-I topology. Pssm-ID: 176036 [Multi-domain] Cd Length: 123 Bit Score: 41.47 E-value: 1.87e-04
|
||||||||||||
| EFh_PI-PLCbeta4 | cd16211 | EF-hand motif found in phosphoinositide phospholipase C beta 4 (PI-PLC-beta4); PI-PLC-beta4, ... |
9-40 | 2.18e-04 | ||||||||
EF-hand motif found in phosphoinositide phospholipase C beta 4 (PI-PLC-beta4); PI-PLC-beta4, also termed 1-phosphatidylinositol 4,5-bisphosphate phosphodiesterase beta-4, or phospholipase C-beta-4 (PLC-beta4), is expressed in high concentrations in cerebellar Purkinje and granule cells, the median geniculate body, and the lateral geniculate nucleus. It may play a critical role in linking anxiety behaviors and theta rhythm heterogeneity. PI-PLC-beta4 is activated by the heterotrimeric G protein alpha q subunits through their C2 domain and long C-terminal extension. It contributes to generate cell-specific Ca2+ signals evoked by G protein-coupled receptor stimulation. PI-PLC-beta4 functions as a downstream signaling molecule of type 1 metabotropic glutamate receptors (mGluR1s). The thalamic mGluR1-PI-PLC-beta4 cascade is essential for formalin-induced inflammatory pain by regulating the response of ventral posterolateral thalamic nucleus (VPL) neurons. Moreover, PI-PLC-beta4 is essential for long-term depression (LTD) in the rostral cerebellum, which may be required for the acquisition of the conditioned eyeblink response. Besides, PI-PLC-beta4 may play an important role in maintenance of the status epilepticus. The mutations of PI-PLC-beta4 has been identified as the major cause of autosomal dominant auriculocondylar syndrome (ACS). PI-PLC-beta4 contains a core set of domains, including an N-terminal pleckstrin homology (PH) domain, four atypical EF-hand motifs, a PLC catalytic core, and a single C2 domain. Besides, it has a unique C-terminal coiled-coil (CT) domain necessary for homodimerization. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. Pssm-ID: 320041 Cd Length: 153 Bit Score: 42.02 E-value: 2.18e-04
|
||||||||||||
| EFh_NorpA_like | cd16212 | EF-hand motif found in Drosophila melanogaster No receptor potential A protein (NorpA) and ... |
10-40 | 2.88e-04 | ||||||||
EF-hand motif found in Drosophila melanogaster No receptor potential A protein (NorpA) and similar proteins; NorpA, also termed 1-phosphatidylinositol 4,5-bisphosphate phosphodiesterase, is an eye-specific phosphoinositide phospholipase C (PI-PLC) encoded by norpA gene in Drosophila. It is expressed predominantly in photoreceptors and plays an essential role in the phototransduction pathway of Drosophila. A mutation within the norpA gene can render the fly blind without affecting any of the obvious structures of the eye. Like beta-class of vertebrate PI-PLCs, NorpA contains an N-terminal pleckstrin homology (PH) domain, four atypical EF-hand motifs, a PLC catalytic core, and a single C2 domain. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. Pssm-ID: 320042 [Multi-domain] Cd Length: 153 Bit Score: 41.38 E-value: 2.88e-04
|
||||||||||||
| EFh_PI-PLCbeta | cd16200 | EF-hand motif found in metazoan phosphoinositide-specific phospholipase C (PI-PLC)-beta ... |
9-40 | 3.91e-04 | ||||||||
EF-hand motif found in metazoan phosphoinositide-specific phospholipase C (PI-PLC)-beta isozymes; PI-PLC-beta isozymes represent a class of metazoan PI-PLCs that hydrolyze the membrane lipid phosphatidylinositol 4,5-bisphosphate (PIP2) to propagate diverse intracellular responses that underlie the physiological action of many hormones, neurotransmitters, and growth factors (EC 3.1.4.11). They have been implicated in numerous processes relevant to central nervous system (CNS), including chemotaxis, cardiovascular function, neuronal signaling, and opioid sensitivity. Like other PI-PLC isozymes, PI-PLC-beta isozymes contain a core set of domains, including an N-terminal pleckstrin homology (PH) domain, four atypical EF-hand motifs, a PLC catalytic core, and a single C2 domain. Besides, they have a unique C-terminal coiled-coil (CT) domain necessary for homodimerization. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. There are four PI-PLC-beta isozymes (1-4). PI-PLC-beta1 and PI-PLC-beta3 are expressed in a wide range of tissues and cell types, whereas PI-PLC-beta2 and PI-PLC-beta4 have been found only in hematopoietic and neuronal tissues, respectively. All PI-PLC-beta isozymes are activated by the heterotrimeric G protein alpha subunits of the Gq class through their C2 domain and long C-terminal extension. They are GTPase-activating proteins (GAPs) for these G alpha(q) proteins. PI-PLC-beta2 and PI-PLC-beta3 can also be activated by beta-gamma subunits of the G alpha(i/o) family of heterotrimeric G proteins and the small GTPases such as Rac and Cdc42. This family also includes two invertebrate homologs of PI-PLC-beta, PLC21 from cephalopod retina and No receptor potential A protein (NorpA) from Drosophila melanogaster. Pssm-ID: 320030 [Multi-domain] Cd Length: 153 Bit Score: 41.08 E-value: 3.91e-04
|
||||||||||||
| EFh_PI-PLCdelta1 | cd16217 | EF-hand motif found in phosphoinositide phospholipase C delta 1 (PI-PLC-delta1); PI-PLC-delta1, ... |
10-40 | 4.43e-04 | ||||||||
EF-hand motif found in phosphoinositide phospholipase C delta 1 (PI-PLC-delta1); PI-PLC-delta1, also termed 1-phosphatidylinositol 4,5-bisphosphate phosphodiesterase delta-1 (PLCD1), or phospholipase C-III (PLC-III), or phospholipase C-delta-1 (PLC-delta-1), is present in high abundancy in the brain, heart, lung, skeletal muscle and testis. It is activated by high calcium levels generated by other PI-PLC family members, and therefore functions as a calcium amplifier within the cell. PI-PLC-delta1 is required for maintenance of homeostasis in skin and metabolic tissues. Moreover, it is essential in trophoblasts for placental development. Simultaneous loss of PI-PLC-delta1 may cause placental vascular defects, leading to embryonic lethality. PI-PLC-delta1 can be positively or negatively regulated by several binding partners, including p122/Rho GTPase activating protein (RhoGAP), Gha/Transglutaminase II, RalA, and calmodulin. It is involved in Alzheimer's disease and hypertension. Furthermore, PI-PLC-delta1 regulates cell proliferation and cell-cycle progression from G1- to S-phase by control of cyclin E-CDK2 activity and p27 levels. It can be activated by alpha1-adrenoreceptors (AR) in a calcium-dependent manner and may be important for G protein-coupled receptors (GPCR) responses in vascular smooth muscle (VSM). PI-PLC-delta1 may also be involved in noradrenaline (NA)-induced phosphatidylinositol-4,5-bisphosphate (PIP2) hydrolysis and modulate sustained contraction of mesenteric small arteries. In addition, it inhibits thermogenesis and induces lipid accumulation, and therefore contributes to the development of obesity. PI-PLC-delta1 contains a core set of domains, including an N-terminal pleckstrin homology (PH) domain, four atypical EF-hand motifs, a PLC catalytic core, and a single C-terminal C2 domain. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. PI-PLC-delta1 can regulate the binding of PH domain to PIP2 in a Ca2+-dependent manner through its functionally important EF-hand domains. In addition, PI-PLC-delta1 possesses a classical leucine-rich nuclear export sequence (NES) located in the EF hand motifs, as well as a nuclear localization signal within its linker region, both of which may be responsible for translocating PI-PLC-delta1 into and out of the cell nucleus. Pssm-ID: 320047 [Multi-domain] Cd Length: 139 Bit Score: 40.88 E-value: 4.43e-04
|
||||||||||||
| C2B_Ferlin | cd04011 | C2 domain second repeat in Ferlin; Ferlins are involved in vesicle fusion events. Ferlins and ... |
414-503 | 5.17e-04 | ||||||||
C2 domain second repeat in Ferlin; Ferlins are involved in vesicle fusion events. Ferlins and other proteins, such as Synaptotagmins, are implicated in facilitating the fusion process when cell membranes fuse together. There are six known human Ferlins: Dysferlin (Fer1L1), Otoferlin (Fer1L2), Myoferlin (Fer1L3), Fer1L4, Fer1L5, and Fer1L6. Defects in these genes can lead to a wide range of diseases including muscular dystrophy (dysferlin), deafness (otoferlin), and infertility (fer-1, fertilization factor-1). Structurally they have 6 tandem C2 domains, designated as (C2A-C2F) and a single C-terminal transmembrane domain, though there is a new study that disputes this and claims that there are actually 7 tandem C2 domains with another C2 domain inserted between C2D and C2E. In a subset of them (Dysferlin, Myoferlin, and Fer1) there is an additional conserved domain called DysF. C2 domains fold into an 8-standed beta-sandwich that can adopt 2 structural arrangements: Type I and Type II, distinguished by a circular permutation involving their N- and C-terminal beta strands. Many C2 domains are Ca2+-dependent membrane-targeting modules that bind a wide variety of substances including bind phospholipids, inositol polyphosphates, and intracellular proteins. Most C2 domain proteins are either signal transduction enzymes that contain a single C2 domain, such as protein kinase C, or membrane trafficking proteins which contain at least two C2 domains, such as synaptotagmin 1. However, there are a few exceptions to this including RIM isoforms and some splice variants of piccolo/aczonin and intersectin which only have a single C2 domain. C2 domains with a calcium binding region have negatively charged residues, primarily aspartates, that serve as ligands for calcium ions. This cd contains the second C2 repeat, C2B, and has a type-II topology. Pssm-ID: 175978 [Multi-domain] Cd Length: 111 Bit Score: 39.87 E-value: 5.17e-04
|
||||||||||||
| C2B_Munc13-like | cd04009 | C2 domain second repeat in Munc13 (mammalian uncoordinated)-like proteins; C2-like domains are ... |
404-497 | 5.48e-04 | ||||||||
C2 domain second repeat in Munc13 (mammalian uncoordinated)-like proteins; C2-like domains are thought to be involved in phospholipid binding in a Ca2+ independent manner in both Unc13 and Munc13. Caenorabditis elegans Unc13 has a central domain with sequence similarity to PKC, which includes C1 and C2-related domains. Unc13 binds phorbol esters and DAG with high affinity in a phospholipid manner. Mutations in Unc13 results in abnormal neuronal connections and impairment in cholinergic neurotransmission in the nematode. Munc13 is the mammalian homolog which are expressed in the brain. There are 3 isoforms (Munc13-1, -2, -3) and are thought to play a role in neurotransmitter release and are hypothesized to be high-affinity receptors for phorbol esters. Unc13 and Munc13 contain both C1 and C2 domains. There are two C2 related domains present, one central and one at the carboxyl end. Munc13-1 contains a third C2-like domain. Munc13 interacts with syntaxin, synaptobrevin, and synaptotagmin suggesting a role for these as scaffolding proteins. C2 domains fold into an 8-standed beta-sandwich that can adopt 2 structural arrangements: Type I and Type II, distinguished by a circular permutation involving their N- and C-terminal beta strands. Many C2 domains are Ca2+-dependent membrane-targeting modules that bind a wide variety of substances including bind phospholipids, inositol polyphosphates, and intracellular proteins. Most C2 domain proteins are either signal transduction enzymes that contain a single C2 domain, such as protein kinase C, or membrane trafficking proteins which contain at least two C2 domains, such as synaptotagmin 1. However, there are a few exceptions to this including RIM isoforms and some splice variants of piccolo/aczonin and intersectin which only have a single C2 domain. C2 domains with a calcium binding region have negatively charged residues, primarily aspartates, that serve as ligands for calcium ions. This cd contains the third C2 repeat, C2C, and has a type-II topology. Pssm-ID: 175976 [Multi-domain] Cd Length: 133 Bit Score: 40.30 E-value: 5.48e-04
|
||||||||||||
| EFh_PI-PLCepsilon | cd16203 | EF-hand motif found in phosphoinositide phospholipase C epsilon 1 (PI-PLC-epsilon1); ... |
1-36 | 6.56e-04 | ||||||||
EF-hand motif found in phosphoinositide phospholipase C epsilon 1 (PI-PLC-epsilon1); PI-PLC-epsilon1, also termed 1-phosphatidylinositol 4,5-bisphosphate phosphodiesterase epsilon-1, or pancreas-enriched phospholipase C, or phospholipase C-epsilon-1 (PLC-epsilon-1), is dominant in connective tissues and brain. It has been implicated in carcinogenesis, such as in bladder and intestinal tumor, oesophageal squamous cell carcinoma, gastric adenocarcinoma, murine skin cancer, head and neck cancer. PI-PLC-epsilon1 contains an N-terminal CDC25 homology domain with a guanyl-nucleotide exchange factor (GFF) activity, a pleckstrin homology (PH) domain, four atypical EF-hand motifs, a PLC catalytic core domain, a C2 domain, and at least one and perhaps two C-terminal predicted RA (Ras association) domains that are implicated in the binding of small GTPases, such as Ras or Rap, from the Ras family. The PLC catalytic core domain is a TIM barrel with two highly conserved regions (X and Y) split by a highly degenerate linker sequence. There is only one PI-PLC-epsilon isozyme. It is directly activated by G alpha(12/13), G beta gamma, and activated members of Ras and Rho small GTPases. Pssm-ID: 320033 Cd Length: 174 Bit Score: 40.77 E-value: 6.56e-04
|
||||||||||||
| C2B_SLP_1-2-3-4 | cd04020 | C2 domain second repeat present in Synaptotagmin-like proteins 1-4; All Slp members basically ... |
412-497 | 7.67e-04 | ||||||||
C2 domain second repeat present in Synaptotagmin-like proteins 1-4; All Slp members basically share an N-terminal Slp homology domain (SHD) and C-terminal tandem C2 domains (named the C2A domain and the C2B domain) with the SHD and C2 domains being separated by a linker sequence of various length. Slp1/JFC1 and Slp2/exophilin 4 promote granule docking to the plasma membrane. Additionally, their C2A domains are both Ca2+ independent, unlike the case in Slp3 and Slp4/granuphilin in which their C2A domains are Ca2+ dependent. It is thought that SHD (except for the Slp4-SHD) functions as a specific Rab27A/B-binding domain. In addition to Slps, rabphilin, Noc2, and Munc13-4 also function as Rab27-binding proteins. It has been demonstrated that Slp3 and Slp4/granuphilin promote dense-core vesicle exocytosis. C2 domains fold into an 8-standed beta-sandwich that can adopt 2 structural arrangements: Type I and Type II, distinguished by a circular permutation involving their N- and C-terminal beta strands. Many C2 domains are Ca2+-dependent membrane-targeting modules that bind a wide variety of substances including bind phospholipids, inositol polyphosphates, and intracellular proteins. Most C2 domain proteins are either signal transduction enzymes that contain a single C2 domain, such as protein kinase C, or membrane trafficking proteins which contain at least two C2 domains, such as synaptotagmin 1. However, there are a few exceptions to this including RIM isoforms and some splice variants of piccolo/aczonin and intersectin which only have a single C2 domain. C2 domains with a calcium binding region have negatively charged residues, primarily aspartates, that serve as ligands for calcium ions. This cd contains the second C2 repeat, C2B, and has a type-I topology. Pssm-ID: 175987 [Multi-domain] Cd Length: 162 Bit Score: 40.38 E-value: 7.67e-04
|
||||||||||||
| C2_SRC2_like | cd04051 | C2 domain present in Soybean genes Regulated by Cold 2 (SRC2)-like proteins; SRC2 production ... |
411-503 | 1.41e-03 | ||||||||
C2 domain present in Soybean genes Regulated by Cold 2 (SRC2)-like proteins; SRC2 production is a response to pathogen infiltration. The initial response of increased Ca2+ concentrations are coupled to downstream signal transduction pathways via calcium binding proteins. SRC2 contains a single C2 domain which localizes to the plasma membrane and is involved in Ca2+ dependent protein binding. C2 domains fold into an 8-standed beta-sandwich that can adopt 2 structural arrangements: Type I and Type II, distinguished by a circular permutation involving their N- and C-terminal beta strands. Many C2 domains are Ca2+-dependent membrane-targeting modules that bind a wide variety of substances including bind phospholipids, inositol polyphosphates, and intracellular proteins. Most C2 domain proteins are either signal transduction enzymes that contain a single C2 domain, such as protein kinase C, or membrane trafficking proteins which contain at least two C2 domains, such as synaptotagmin 1. However, there are a few exceptions to this including RIM isoforms and some splice variants of piccolo/aczonin and intersectin which only have a single C2 domain. C2 domains with a calcium binding region have negatively charged residues, primarily aspartates, that serve as ligands for calcium ions. Pssm-ID: 176016 [Multi-domain] Cd Length: 125 Bit Score: 38.75 E-value: 1.41e-03
|
||||||||||||
| C2_ArfGAP | cd04038 | C2 domain present in Arf GTPase Activating Proteins (GAP); ArfGAP is a GTPase activating ... |
412-478 | 1.59e-03 | ||||||||
C2 domain present in Arf GTPase Activating Proteins (GAP); ArfGAP is a GTPase activating protein which regulates the ADP ribosylation factor Arf, a member of the Ras superfamily of GTP-binding proteins. The GTP-bound form of Arf is involved in Golgi morphology and is involved in recruiting coat proteins. ArfGAP is responsible for the GDP-bound form of Arf which is necessary for uncoating the membrane and allowing the Golgi to fuse with an acceptor compartment. These proteins contain an N-terminal ArfGAP domain containing the characteristic zinc finger motif (Cys-x2-Cys-x(16,17)-x2-Cys) and C-terminal C2 domain. C2 domains were first identified in Protein Kinase C (PKC). C2 domains fold into an 8-standed beta-sandwich that can adopt 2 structural arrangements: Type I and Type II, distinguished by a circular permutation involving their N- and C-terminal beta strands. Many C2 domains are Ca2+-dependent membrane-targeting modules that bind a wide variety of substances including bind phospholipids, inositol polyphosphates, and intracellular proteins. Most C2 domain proteins are either signal transduction enzymes that contain a single C2 domain, such as protein kinase C, or membrane trafficking proteins which contain at least two C2 domains, such as synaptotagmin 1. However, there are a few exceptions to this including RIM isoforms and some splice variants of piccolo/aczonin and intersectin which only have a single C2 domain. C2 domains with a calcium binding region have negatively charged residues, primarily aspartates, that serve as ligands for calcium ions. Pssm-ID: 176003 [Multi-domain] Cd Length: 145 Bit Score: 39.23 E-value: 1.59e-03
|
||||||||||||
| EFh_PRIP2 | cd16223 | EF-hand motif found in phospholipase C-related but catalytically inactive protein 2 (PRIP-2); ... |
8-41 | 3.55e-03 | ||||||||
EF-hand motif found in phospholipase C-related but catalytically inactive protein 2 (PRIP-2); PRIP-2, also termed phospholipase C-L2, or phospholipase C-epsilon-2 (PLC-epsilon-2), or inactive phospholipase C-like protein 2 (PLC-L2), is a novel inositol 1,4,5-trisphosphate (InsP3) binding protein that exhibits a relatively ubiquitous expression. It functions as a novel negative regulator of B-cell receptor (BCR) signaling and immune responses. PRIP-2 has a primary structure and domain architecture, incorporating a pleckstrin homology (PH) domain, four atypical EF-hand motifs, a PLC catalytic core domain with highly conserved X- and Y-regions split by a linker sequence, and a C-terminal C2 domain, similar to phosphoinositide-specific phospholipases C (PI-PLC, EC 3.1.4.11)-delta isoforms. Due to replacement of critical catalytic residues, PRIP-2 does not have PLC enzymatic activity. Pssm-ID: 320053 [Multi-domain] Cd Length: 144 Bit Score: 38.35 E-value: 3.55e-03
|
||||||||||||
| Blast search parameters | ||||
|
