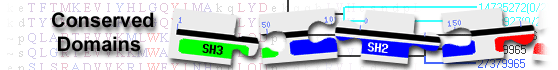


netrin receptor UNC5D isoform X1 [Mus musculus]
Protein Classification

immunoglobulin domain-containing protein( domain architecture ID 11668633)
immunoglobulin (Ig) domain-containing protein adopts a fold comprised of a sandwich of two beta sheets and may function in cell adhesion and pattern recognition; similar to Drosophila melanogaster DIP/Dpr cell recognition proteins, which are members of the Wirin family of IgSF proteins with neuronal wiring functions, and human IgLON proteins, a family of cell adhesion molecules
List of domain hits
| Name | Accession | Description | Interval | E-value | |||
| UPA | pfam17217 | UPA domain; The UPA domain is conserved in UNC5, PIDD, and Ankyrins. It has a beta sandwich ... |
685-824 | 1.23e-82 | |||
UPA domain; The UPA domain is conserved in UNC5, PIDD, and Ankyrins. It has a beta sandwich structure. : Pssm-ID: 465384 Cd Length: 140 Bit Score: 262.68 E-value: 1.23e-82
|
|||||||
| Death_UNC5D | cd08801 | Death domain found in Uncoordinated-5D; Death Domain (DD) found in Uncoordinated-5D (UNC5D). ... |
851-948 | 1.14e-64 | |||
Death domain found in Uncoordinated-5D; Death Domain (DD) found in Uncoordinated-5D (UNC5D). UNC5D is part of the UNC-5 homolog family. It is a receptor for the secreted netrin-1 and plays a role in axonal guidance, angiogenesis, and apoptosis. UNC5 proteins are transmembrane proteins with an extracellular domain consisting of two immunoglobulin repeats, two thrombospondin type-I modules and an intracellular region containing a ZU-5 domain, UPA domain and a DD. In general, DDs are protein-protein interaction domains found in a variety of domain architectures. Their common feature is that they form homodimers by self-association or heterodimers by associating with other members of the DD superfamily including CARD (Caspase activation and recruitment domain), DED (Death Effector Domain), and PYRIN. They serve as adaptors in signaling pathways and can recruit other proteins into signaling complexes. : Pssm-ID: 176779 Cd Length: 98 Bit Score: 212.22 E-value: 1.14e-64
|
|||||||
| ZU5 | pfam00791 | ZU5 domain; Domain present in ZO-1 and Unc5-like netrin receptors Domain of unknown function. |
542-637 | 3.47e-40 | |||
ZU5 domain; Domain present in ZO-1 and Unc5-like netrin receptors Domain of unknown function. : Pssm-ID: 459941 Cd Length: 97 Bit Score: 143.05 E-value: 3.47e-40
|
|||||||
| Ig super family | cl11960 | Immunoglobulin domain; The members here are composed of the immunoglobulin (Ig) domain found ... |
161-247 | 2.68e-18 | |||
Immunoglobulin domain; The members here are composed of the immunoglobulin (Ig) domain found in the Ig superfamily. The Ig superfamily is a heterogenous group of proteins, built on a common fold comprised of a sandwich of two beta sheets. Members of this group are components of immunoglobulin, neuroglia, cell surface glycoproteins, including T-cell receptors, CD2, CD4, CD8, and membrane glycoproteins, including butyrophilin and chondroitin sulfate proteoglycan core protein. A predominant feature of most Ig domains is a disulfide bridge connecting the two beta-sheets with a tryptophan residue packed against the disulfide bond. Ig superfamily (IgSF) domains can be divided into 4 main classes based on their structures and sequences: the Variable (V), Constant 1 (C1), Constant 2 (C2), and Intermediate (I) sets. Typically, the V-set domains have A, B, E, and D strands in one sheet and A', G, F, C, C' and C" in the other. The structures in C1-set are smaller than those in the V-set; they have one beta sheet that is formed by strands A, B, E, and D and the other by strands G, F, C, and C'. Moreover, a C1-set Ig domain contains a short C' strand (three residues) and lacks A' and C" strand. Unlike other Ig domain sets, C2-set structures do not have a D strand. Like the V-set Ig domains, members of the I-set have a discontinuous A strand, but lack a C" strand. The actual alignment was detected with superfamily member cd05724: Pssm-ID: 472250 [Multi-domain] Cd Length: 87 Bit Score: 80.52 E-value: 2.68e-18
|
|||||||
| TSP1 | smart00209 | Thrombospondin type 1 repeats; Type 1 repeats in thrombospondin-1 bind and activate TGF-beta. |
253-304 | 1.96e-15 | |||
Thrombospondin type 1 repeats; Type 1 repeats in thrombospondin-1 bind and activate TGF-beta. : Pssm-ID: 214559 [Multi-domain] Cd Length: 53 Bit Score: 71.08 E-value: 1.96e-15
|
|||||||
| TSP1 | smart00209 | Thrombospondin type 1 repeats; Type 1 repeats in thrombospondin-1 bind and activate TGF-beta. |
309-357 | 4.46e-10 | |||
Thrombospondin type 1 repeats; Type 1 repeats in thrombospondin-1 bind and activate TGF-beta. : Pssm-ID: 214559 [Multi-domain] Cd Length: 53 Bit Score: 56.06 E-value: 4.46e-10
|
|||||||
| Ig super family | cl11960 | Immunoglobulin domain; The members here are composed of the immunoglobulin (Ig) domain found ... |
52-153 | 2.17e-03 | |||
Immunoglobulin domain; The members here are composed of the immunoglobulin (Ig) domain found in the Ig superfamily. The Ig superfamily is a heterogenous group of proteins, built on a common fold comprised of a sandwich of two beta sheets. Members of this group are components of immunoglobulin, neuroglia, cell surface glycoproteins, including T-cell receptors, CD2, CD4, CD8, and membrane glycoproteins, including butyrophilin and chondroitin sulfate proteoglycan core protein. A predominant feature of most Ig domains is a disulfide bridge connecting the two beta-sheets with a tryptophan residue packed against the disulfide bond. Ig superfamily (IgSF) domains can be divided into 4 main classes based on their structures and sequences: the Variable (V), Constant 1 (C1), Constant 2 (C2), and Intermediate (I) sets. Typically, the V-set domains have A, B, E, and D strands in one sheet and A', G, F, C, C' and C" in the other. The structures in C1-set are smaller than those in the V-set; they have one beta sheet that is formed by strands A, B, E, and D and the other by strands G, F, C, and C'. Moreover, a C1-set Ig domain contains a short C' strand (three residues) and lacks A' and C" strand. Unlike other Ig domain sets, C2-set structures do not have a D strand. Like the V-set Ig domains, members of the I-set have a discontinuous A strand, but lack a C" strand. The actual alignment was detected with superfamily member cd07693: Pssm-ID: 472250 [Multi-domain] Cd Length: 99 Bit Score: 38.30 E-value: 2.17e-03
|
|||||||
| Name | Accession | Description | Interval | E-value | |||
| UPA | pfam17217 | UPA domain; The UPA domain is conserved in UNC5, PIDD, and Ankyrins. It has a beta sandwich ... |
685-824 | 1.23e-82 | |||
UPA domain; The UPA domain is conserved in UNC5, PIDD, and Ankyrins. It has a beta sandwich structure. Pssm-ID: 465384 Cd Length: 140 Bit Score: 262.68 E-value: 1.23e-82
|
|||||||
| Death_UNC5D | cd08801 | Death domain found in Uncoordinated-5D; Death Domain (DD) found in Uncoordinated-5D (UNC5D). ... |
851-948 | 1.14e-64 | |||
Death domain found in Uncoordinated-5D; Death Domain (DD) found in Uncoordinated-5D (UNC5D). UNC5D is part of the UNC-5 homolog family. It is a receptor for the secreted netrin-1 and plays a role in axonal guidance, angiogenesis, and apoptosis. UNC5 proteins are transmembrane proteins with an extracellular domain consisting of two immunoglobulin repeats, two thrombospondin type-I modules and an intracellular region containing a ZU-5 domain, UPA domain and a DD. In general, DDs are protein-protein interaction domains found in a variety of domain architectures. Their common feature is that they form homodimers by self-association or heterodimers by associating with other members of the DD superfamily including CARD (Caspase activation and recruitment domain), DED (Death Effector Domain), and PYRIN. They serve as adaptors in signaling pathways and can recruit other proteins into signaling complexes. Pssm-ID: 176779 Cd Length: 98 Bit Score: 212.22 E-value: 1.14e-64
|
|||||||
| ZU5 | pfam00791 | ZU5 domain; Domain present in ZO-1 and Unc5-like netrin receptors Domain of unknown function. |
542-637 | 3.47e-40 | |||
ZU5 domain; Domain present in ZO-1 and Unc5-like netrin receptors Domain of unknown function. Pssm-ID: 459941 Cd Length: 97 Bit Score: 143.05 E-value: 3.47e-40
|
|||||||
| ZU5 | smart00218 | Domain present in ZO-1 and Unc5-like netrin receptors; Domain of unknown function. |
544-640 | 2.89e-23 | |||
Domain present in ZO-1 and Unc5-like netrin receptors; Domain of unknown function. Pssm-ID: 128514 Cd Length: 104 Bit Score: 95.11 E-value: 2.89e-23
|
|||||||
| IgI_2_Robo | cd05724 | Second immunoglobulin (Ig)-like domain in Robo (roundabout) receptors; member of the I-set of ... |
161-247 | 2.68e-18 | |||
Second immunoglobulin (Ig)-like domain in Robo (roundabout) receptors; member of the I-set of Ig superfamily (IgSF) domains; The members here are composed of the second immunoglobulin (Ig)-like domain in Robo (roundabout) receptors. Robo receptors play a role in the development of the central nervous system (CNS), and are receptors of the Slit protein. Slit is a repellant secreted by the neural cells in the midline. Slit acts through Robo to prevent most neurons from crossing the midline from either side. Three mammalian Robo homologs (Robo1, Robo2, and Robo3), and three mammalian Slit homologs (Slit-1,Slit-2, Slit-3), have been identified. Commissural axons, which cross the midline, express low levels of Robo; longitudinal axons, which avoid the midline, express high levels of Robo. Robo1, Robo2, and Robo3 are expressed by commissural neurons in the vertebrate spinal cord and Slit-1, Slit-2, Slit-3 are expressed at the ventral midline. Robo-3 is a divergent member of the Robo family which instead of being a positive regulator of Slit responsiveness, antagonizes Slit responsiveness in precrossing axons. The Slit-Robo interaction is mediated by the second leucine-rich repeat (LRR) domain of Slit and the two N-terminal Ig domains of Robo, Ig1 and Ig2. The primary Robo binding site for Slit-2 has been shown by surface plasmon resonance experiments and mutational analysis to be the Ig1 domain, while the Ig2 domain has been proposed to harbor a weak secondary binding site. This group belongs to the I-set of IgSF domains, having A-B-E-D strands in one beta-sheet and A'-G-F-C-C' in the other. Like the V-set Ig domains, members of the I-set have a discontinuous A strand, but lack a C" strand. Pssm-ID: 409389 [Multi-domain] Cd Length: 87 Bit Score: 80.52 E-value: 2.68e-18
|
|||||||
| Death | pfam00531 | Death domain; |
856-927 | 5.34e-16 | |||
Death domain; Pssm-ID: 459845 [Multi-domain] Cd Length: 86 Bit Score: 73.94 E-value: 5.34e-16
|
|||||||
| TSP1 | smart00209 | Thrombospondin type 1 repeats; Type 1 repeats in thrombospondin-1 bind and activate TGF-beta. |
253-304 | 1.96e-15 | |||
Thrombospondin type 1 repeats; Type 1 repeats in thrombospondin-1 bind and activate TGF-beta. Pssm-ID: 214559 [Multi-domain] Cd Length: 53 Bit Score: 71.08 E-value: 1.96e-15
|
|||||||
| I-set | pfam07679 | Immunoglobulin I-set domain; |
159-247 | 7.34e-11 | |||
Immunoglobulin I-set domain; Pssm-ID: 400151 [Multi-domain] Cd Length: 90 Bit Score: 59.19 E-value: 7.34e-11
|
|||||||
| TSP1 | smart00209 | Thrombospondin type 1 repeats; Type 1 repeats in thrombospondin-1 bind and activate TGF-beta. |
309-357 | 4.46e-10 | |||
Thrombospondin type 1 repeats; Type 1 repeats in thrombospondin-1 bind and activate TGF-beta. Pssm-ID: 214559 [Multi-domain] Cd Length: 53 Bit Score: 56.06 E-value: 4.46e-10
|
|||||||
| TSP_1 | pfam00090 | Thrombospondin type 1 domain; |
254-301 | 1.04e-08 | |||
Thrombospondin type 1 domain; Pssm-ID: 459668 [Multi-domain] Cd Length: 49 Bit Score: 52.04 E-value: 1.04e-08
|
|||||||
| DEATH | smart00005 | DEATH domain, found in proteins involved in cell death (apoptosis); Alpha-helical domain ... |
855-927 | 1.96e-08 | |||
DEATH domain, found in proteins involved in cell death (apoptosis); Alpha-helical domain present in a variety of proteins with apoptotic functions. Some (but not all) of these domains form homotypic and heterotypic dimers. Pssm-ID: 214467 [Multi-domain] Cd Length: 88 Bit Score: 52.41 E-value: 1.96e-08
|
|||||||
| IG_like | smart00410 | Immunoglobulin like; IG domains that cannot be classified into one of IGv1, IGc1, IGc2, IG. |
163-249 | 3.50e-08 | |||
Immunoglobulin like; IG domains that cannot be classified into one of IGv1, IGc1, IGc2, IG. Pssm-ID: 214653 [Multi-domain] Cd Length: 85 Bit Score: 51.74 E-value: 3.50e-08
|
|||||||
| TSP_1 | pfam00090 | Thrombospondin type 1 domain; |
311-357 | 5.45e-05 | |||
Thrombospondin type 1 domain; Pssm-ID: 459668 [Multi-domain] Cd Length: 49 Bit Score: 41.25 E-value: 5.45e-05
|
|||||||
| IgC_1_Robo | cd07693 | First immunoglobulin (Ig)-like constant domain in Robo (roundabout) receptors, and similar ... |
52-153 | 2.17e-03 | |||
First immunoglobulin (Ig)-like constant domain in Robo (roundabout) receptors, and similar domains; The members here are composed of the first immunoglobulin (Ig)-like domain in Roundabout (Robo) receptors. Robo receptors play a role in the development of the central nervous system (CNS), and are receptors of Slit protein. Slit is a repellant secreted by the neural cells in the midline. Slit acts through Robo to prevent most neurons from crossing the midline from either side. Three mammalian Robo homologs (Robo1, Robo2, and Robo3), and three mammalian Slit homologs (Slit1, Slit2, Slit3), have been identified. Commissural axons, which cross the midline, express low levels of Robo; longitudinal axons, which avoid the midline, express high levels of Robo. Robo1, Robo2, and Robo3 are expressed by commissural neurons in the vertebrate spinal cord and Slit1, Slit2,and Slit3 are expressed at the ventral midline. Robo3 is a divergent member of the Robo family which instead of being a positive regulator of Slit responsiveness, antagonizes Slit responsiveness in precrossing axons. The Slit-Robo interaction is mediated by the second leucine-rich repeat (LRR) domain of Slit and the two N-terminal Ig domains of Robo, Ig1 and Ig2. The primary Robo binding site for Slit2 has been shown by surface plasmon resonance experiments and mutational analysis to be is the Ig1 domain, while the Ig2 domain has been proposed to harbor a weak secondary binding site. Pssm-ID: 409490 [Multi-domain] Cd Length: 99 Bit Score: 38.30 E-value: 2.17e-03
|
|||||||
| Name | Accession | Description | Interval | E-value | |||
| UPA | pfam17217 | UPA domain; The UPA domain is conserved in UNC5, PIDD, and Ankyrins. It has a beta sandwich ... |
685-824 | 1.23e-82 | |||
UPA domain; The UPA domain is conserved in UNC5, PIDD, and Ankyrins. It has a beta sandwich structure. Pssm-ID: 465384 Cd Length: 140 Bit Score: 262.68 E-value: 1.23e-82
|
|||||||
| Death_UNC5D | cd08801 | Death domain found in Uncoordinated-5D; Death Domain (DD) found in Uncoordinated-5D (UNC5D). ... |
851-948 | 1.14e-64 | |||
Death domain found in Uncoordinated-5D; Death Domain (DD) found in Uncoordinated-5D (UNC5D). UNC5D is part of the UNC-5 homolog family. It is a receptor for the secreted netrin-1 and plays a role in axonal guidance, angiogenesis, and apoptosis. UNC5 proteins are transmembrane proteins with an extracellular domain consisting of two immunoglobulin repeats, two thrombospondin type-I modules and an intracellular region containing a ZU-5 domain, UPA domain and a DD. In general, DDs are protein-protein interaction domains found in a variety of domain architectures. Their common feature is that they form homodimers by self-association or heterodimers by associating with other members of the DD superfamily including CARD (Caspase activation and recruitment domain), DED (Death Effector Domain), and PYRIN. They serve as adaptors in signaling pathways and can recruit other proteins into signaling complexes. Pssm-ID: 176779 Cd Length: 98 Bit Score: 212.22 E-value: 1.14e-64
|
|||||||
| ZU5 | pfam00791 | ZU5 domain; Domain present in ZO-1 and Unc5-like netrin receptors Domain of unknown function. |
542-637 | 3.47e-40 | |||
ZU5 domain; Domain present in ZO-1 and Unc5-like netrin receptors Domain of unknown function. Pssm-ID: 459941 Cd Length: 97 Bit Score: 143.05 E-value: 3.47e-40
|
|||||||
| Death_UNC5-like | cd08781 | Death domain found in Uncoordinated-5 homolog family; Death Domain (DD) found in ... |
851-933 | 4.59e-39 | |||
Death domain found in Uncoordinated-5 homolog family; Death Domain (DD) found in Uncoordinated-5 (UNC-5) homolog family, which includes Unc5A, B, C and D in vertebrates. UNC5 proteins are receptors for secreted netrins (netrin-1, -3 and -4) that are involved in diverse processes like axonal guidance, neuronal migration, blood vessel patterning, and apoptosis. They are transmembrane proteins with an extracellular domain consisting of two immunoglobulin repeats, two thrombospondin type-I modules and an intracellular region containing a ZU-5 domain, UPA domain and a DD. In general, DDs are protein-protein interaction domains found in a variety of domain architectures. Their common feature is that they form homodimers by self-association or heterodimers by associating with other members of the DD superfamily including CARD (Caspase activation and recruitment domain), DED (Death Effector Domain), and PYRIN. They serve as adaptors in signaling pathways and can recruit other proteins into signaling complexes. Pssm-ID: 260051 Cd Length: 83 Bit Score: 139.33 E-value: 4.59e-39
|
|||||||
| Death_UNC5B | cd08802 | Death domain found in Uncoordinated-5B; Death Domain (DD) found in Uncoordinated-5B (UNC5B). ... |
851-934 | 2.02e-33 | |||
Death domain found in Uncoordinated-5B; Death Domain (DD) found in Uncoordinated-5B (UNC5B). UNC5B is part of the UNC-5 homolog family. It is a receptor for the secreted netrin-1 and plays a role in axonal guidance, angiogenesis, and apoptosis. UNC5B signaling is involved in the netrin-1-induced proliferation and migration of renal proximal tubular cells. It is also required for vascular patterning during embryonic development, and its activation inhibits sprouting angiogenesis. UNC5 proteins are transmembrane proteins with an extracellular domain consisting of two immunoglobulin repeats, two thrombospondin type-I modules and an intracellular region containing a ZU-5 domain, UPA domain and a DD. In general, DDs are protein-protein interaction domains found in a variety of domain architectures. Their common feature is that they form homodimers by self-association or heterodimers by associating with other members of the DD superfamily including CARD (Caspase activation and recruitment domain), DED (Death Effector Domain), and PYRIN. They serve as adaptors in signaling pathways and can recruit other proteins into signaling complexes. Pssm-ID: 176780 Cd Length: 84 Bit Score: 123.59 E-value: 2.02e-33
|
|||||||
| Death_UNC5C | cd08799 | Death domain found in Uncoordinated-5C; Death Domain (DD) found in Uncoordinated-5C (UNC5C). ... |
851-931 | 1.13e-27 | |||
Death domain found in Uncoordinated-5C; Death Domain (DD) found in Uncoordinated-5C (UNC5C). UNC5C is part of the UNC-5 homolog family. It is a receptor for the secreted netrin-1 and plays a role in axonal guidance, angiogenesis, and apoptosis. UNC5C plays a critical role in the development of spinal accessory motor neurons. Methylation of the UNC5C gene is associated with early stages of colorectal carcinogenesis. UNC5 proteins are transmembrane proteins with an extracellular domain consisting of two immunoglobulin repeats, two thrombospondin type-I modules and an intracellular region containing a ZU-5 domain, UPA domain and a DD. In general, DDs are protein-protein interaction domains found in a variety of domain architectures. Their common feature is that they form homodimers by self-association or heterodimers by associating with other members of the DD superfamily including CARD (Caspase activation and recruitment domain), DED (Death Effector Domain), and PYRIN. They serve as adaptors in signaling pathways and can recruit other proteins into signaling complexes. Pssm-ID: 260064 Cd Length: 83 Bit Score: 107.02 E-value: 1.13e-27
|
|||||||
| Death_UNC5A | cd08800 | Death domain found in Uncoordinated-5A; Death Domain (DD) found in Uncoordinated-5A (UNC5A). ... |
851-925 | 3.07e-25 | |||
Death domain found in Uncoordinated-5A; Death Domain (DD) found in Uncoordinated-5A (UNC5A). UNC5A is part of the UNC-5 homolog family. It is a receptor for the secreted netrin-1 and plays a critical role in neuronal development and differentiation, as well as axon-guidance. It also plays a role in regulating apoptosis in non-neuronal cells as a downstream target of p53. UNC5 proteins are transmembrane proteins with an extracellular domain consisting of two immunoglobulin repeats, two thrombospondin type-I modules and an intracellular region containing a ZU-5 domain, UPA domain and a DD. In general, DDs are protein-protein interaction domains found in a variety of domain architectures. Their common feature is that they form homodimers by self-association or heterodimers by associating with other members of the DD superfamily including CARD (Caspase activation and recruitment domain), DED (Death Effector Domain), and PYRIN. They serve as adaptors in signaling pathways and can recruit other proteins into signaling complexes. Pssm-ID: 260065 Cd Length: 84 Bit Score: 99.96 E-value: 3.07e-25
|
|||||||
| ZU5 | smart00218 | Domain present in ZO-1 and Unc5-like netrin receptors; Domain of unknown function. |
544-640 | 2.89e-23 | |||
Domain present in ZO-1 and Unc5-like netrin receptors; Domain of unknown function. Pssm-ID: 128514 Cd Length: 104 Bit Score: 95.11 E-value: 2.89e-23
|
|||||||
| IgI_2_Robo | cd05724 | Second immunoglobulin (Ig)-like domain in Robo (roundabout) receptors; member of the I-set of ... |
161-247 | 2.68e-18 | |||
Second immunoglobulin (Ig)-like domain in Robo (roundabout) receptors; member of the I-set of Ig superfamily (IgSF) domains; The members here are composed of the second immunoglobulin (Ig)-like domain in Robo (roundabout) receptors. Robo receptors play a role in the development of the central nervous system (CNS), and are receptors of the Slit protein. Slit is a repellant secreted by the neural cells in the midline. Slit acts through Robo to prevent most neurons from crossing the midline from either side. Three mammalian Robo homologs (Robo1, Robo2, and Robo3), and three mammalian Slit homologs (Slit-1,Slit-2, Slit-3), have been identified. Commissural axons, which cross the midline, express low levels of Robo; longitudinal axons, which avoid the midline, express high levels of Robo. Robo1, Robo2, and Robo3 are expressed by commissural neurons in the vertebrate spinal cord and Slit-1, Slit-2, Slit-3 are expressed at the ventral midline. Robo-3 is a divergent member of the Robo family which instead of being a positive regulator of Slit responsiveness, antagonizes Slit responsiveness in precrossing axons. The Slit-Robo interaction is mediated by the second leucine-rich repeat (LRR) domain of Slit and the two N-terminal Ig domains of Robo, Ig1 and Ig2. The primary Robo binding site for Slit-2 has been shown by surface plasmon resonance experiments and mutational analysis to be the Ig1 domain, while the Ig2 domain has been proposed to harbor a weak secondary binding site. This group belongs to the I-set of IgSF domains, having A-B-E-D strands in one beta-sheet and A'-G-F-C-C' in the other. Like the V-set Ig domains, members of the I-set have a discontinuous A strand, but lack a C" strand. Pssm-ID: 409389 [Multi-domain] Cd Length: 87 Bit Score: 80.52 E-value: 2.68e-18
|
|||||||
| Death | pfam00531 | Death domain; |
856-927 | 5.34e-16 | |||
Death domain; Pssm-ID: 459845 [Multi-domain] Cd Length: 86 Bit Score: 73.94 E-value: 5.34e-16
|
|||||||
| TSP1 | smart00209 | Thrombospondin type 1 repeats; Type 1 repeats in thrombospondin-1 bind and activate TGF-beta. |
253-304 | 1.96e-15 | |||
Thrombospondin type 1 repeats; Type 1 repeats in thrombospondin-1 bind and activate TGF-beta. Pssm-ID: 214559 [Multi-domain] Cd Length: 53 Bit Score: 71.08 E-value: 1.96e-15
|
|||||||
| Ig | cd00096 | Immunoglobulin domain; The members here are composed of the immunoglobulin (Ig) domain found ... |
174-243 | 5.76e-11 | |||
Immunoglobulin domain; The members here are composed of the immunoglobulin (Ig) domain found in the Ig superfamily. The Ig superfamily is a heterogenous group of proteins, built on a common fold comprised of a sandwich of two beta sheets. Members of this group are components of immunoglobulin, neuroglia, cell surface glycoproteins, including T-cell receptors, CD2, CD4, CD8, and membrane glycoproteins, including butyrophilin and chondroitin sulfate proteoglycan core protein. A predominant feature of most Ig domains is a disulfide bridge connecting the two beta-sheets with a tryptophan residue packed against the disulfide bond. Ig superfamily (IgSF) domains can be divided into 4 main classes based on their structures and sequences: the Variable (V), Constant 1 (C1), Constant 2 (C2), and Intermediate (I) sets. Typically, the V-set domains have A, B, E, and D strands in one sheet and A', G, F, C, C' and C" in the other. The structures in C1-set are smaller than those in the V-set; they have one beta sheet that is formed by strands A, B, E, and D and the other by strands G, F, C, and C'. Moreover, a C1-set Ig domain contains a short C' strand (three residues) and lacks A' and C" strand. Unlike other Ig domain sets, C2-set structures do not have a D strand. Like the V-set Ig domains, members of the I-set have a discontinuous A strand, but lack a C" strand. Pssm-ID: 409353 [Multi-domain] Cd Length: 70 Bit Score: 58.88 E-value: 5.76e-11
|
|||||||
| I-set | pfam07679 | Immunoglobulin I-set domain; |
159-247 | 7.34e-11 | |||
Immunoglobulin I-set domain; Pssm-ID: 400151 [Multi-domain] Cd Length: 90 Bit Score: 59.19 E-value: 7.34e-11
|
|||||||
| TSP1 | smart00209 | Thrombospondin type 1 repeats; Type 1 repeats in thrombospondin-1 bind and activate TGF-beta. |
309-357 | 4.46e-10 | |||
Thrombospondin type 1 repeats; Type 1 repeats in thrombospondin-1 bind and activate TGF-beta. Pssm-ID: 214559 [Multi-domain] Cd Length: 53 Bit Score: 56.06 E-value: 4.46e-10
|
|||||||
| IgI_5_Robo | cd20952 | Fifth Ig-like domain of Roundabout (Robo) homolog 1/2, and similar domains; a member of the ... |
159-247 | 8.23e-10 | |||
Fifth Ig-like domain of Roundabout (Robo) homolog 1/2, and similar domains; a member of the I-set of IgSF domains; The members here are composed of the fifth Ig-like domain of Roundabout (Robo) homolog 1/2 and similar domains. Robo receptors play a role in the development of the central nervous system (CNS), and are receptors of Slit protein. Slit is a repellant secreted by the neural cells in the midline. Slit acts through Robo to prevent most neurons from crossing the midline from either side. Three mammalian Robo homologs (Robo1, -2, and -3), and three mammalian Slit homologs (Slit-1,-2, -3), have been identified. Commissural axons, which cross the midline, express low levels of Robo; longitudinal axons, which avoid the midline, express high levels of Robo. Robo1, -2, and -3 are expressed by commissural neurons in the vertebrate spinal cord and Slits 1, -2, -3 are expressed at the ventral midline. Robo-3 is a divergent member of the Robo family which instead of being a positive regulator of slit responsiveness, antagonizes slit responsiveness in precrossing axons. The Slit-Robo interaction is mediated by the second leucine-rich repeat (LRR) domain of Slit and the two N-terminal Ig domains of Robo, Ig1 and Ig2. The primary Robo binding site for Slit2 has been shown by surface plasmon resonance experiments and mutational analysis to be is the Ig1 domain, while the Ig2 domain has been proposed to harbor a weak secondary binding site. The fifth Ig-like domain of Robo 1 and 2 is a member of the I-set Ig domains, having A-B-E-D strands in one beta-sheet and A'-G-F-C-C' in the other. Like the V-set Ig domains, members of the I-set have a discontinuous A strand but lack a C" strand. I-set domains are found in several cell adhesion molecules (such as VCAM, ICAM, and MADCAM), and are also present in numerous other diverse protein families, including several tyrosine-protein kinase receptors Pssm-ID: 409544 [Multi-domain] Cd Length: 87 Bit Score: 56.35 E-value: 8.23e-10
|
|||||||
| IgI_2_Follistatin_like | cd05736 | Second immunoglobulin (Ig)-like domain of a Follistatin-related protein 5, and similar domains; ... |
164-246 | 2.52e-09 | |||
Second immunoglobulin (Ig)-like domain of a Follistatin-related protein 5, and similar domains; member of the I-set of Ig superfamily (IgSF) domains; The members here are composed of the second immunoglobulin (Ig)-like domain found in human Follistatin-related protein 5 (FSTL5) and a follistatin-like molecule encoded by the CNS-related Mahya gene. Mahya genes have been retained in certain Bilaterian branches during evolution. They are conserved in Hymenoptera and Deuterostomes, but are absent from other metazoan species such as fruit fly and nematode. Mahya proteins are secretory, with a follistatin-like domain (Kazal-type serine/threonine protease inhibitor domain and EF-hand calcium-binding domain), two Ig-like domains, and a novel C-terminal domain. Mahya may be involved in learning and memory and in processing of sensory information in Hymenoptera and vertebrates. Follistatin is a secreted, multidomain protein that binds activins with high affinity and antagonizes their signaling. Pssm-ID: 409399 [Multi-domain] Cd Length: 93 Bit Score: 54.96 E-value: 2.52e-09
|
|||||||
| Ig_Pro_neuregulin | cd05750 | Immunoglobulin (Ig)-like domain in neuregulins; The members here are composed of the ... |
174-249 | 3.37e-09 | |||
Immunoglobulin (Ig)-like domain in neuregulins; The members here are composed of the immunoglobulin (Ig)-like domain in neuregulins (NRGs). NRGs are signaling molecules which participate in cell-cell interactions in the nervous system, breast, heart, and other organ systems, and are implicated in the pathology of diseases including schizophrenia, multiple sclerosis, and breast cancer. There are four members of the neuregulin gene family (NRG-1, NRG-2, NRG-3, and NRG-4). The NRG-1 protein, binds to and activates the tyrosine kinases receptors ErbB3 and ErbB4, initiating signaling cascades. The other NRGs proteins bind one or the other or both of these ErbBs. NRG-1 has multiple functions: in the brain it regulates various processes such as radial glia formation and neuronal migration, dendritic development, and expression of neurotransmitters receptors, while in the peripheral nervous system NRG-1 regulates processes such as target cell differentiation, and Schwann cell survival. There are many NRG-1 isoforms which arise from the alternative splicing of mRNA. Less is known of the functions of the other NRGs. NRG-2 and NRG-3 are expressed predominantly in the nervous system. NRG-2 is expressed by motor neurons and terminal Schwann cells, and is concentrated near synaptic sites and may be a signal that regulates synaptic differentiation. NRG-4 has been shown to direct pancreatic islet cell development towards the delta-cell lineage. Pssm-ID: 409408 [Multi-domain] Cd Length: 92 Bit Score: 54.82 E-value: 3.37e-09
|
|||||||
| Ig_3 | pfam13927 | Immunoglobulin domain; This family contains immunoglobulin-like domains. |
163-233 | 4.27e-09 | |||
Immunoglobulin domain; This family contains immunoglobulin-like domains. Pssm-ID: 464046 [Multi-domain] Cd Length: 78 Bit Score: 54.11 E-value: 4.27e-09
|
|||||||
| IgI_2_Titin_Z1z2-like | cd20972 | Second Ig-like domain of the giant muscle protein titin Z1z2 in the sarcomeric Z-disk, and ... |
158-247 | 7.75e-09 | |||
Second Ig-like domain of the giant muscle protein titin Z1z2 in the sarcomeric Z-disk, and similar domains; a member of the I-set of IgSF domains; The members here are composed of the second immunoglobulin (Ig)-like domain of the giant muscle protein titin Z1z2 in the sarcomeric Z-disk and similar proteins. Titin is a key component in the assembly and functioning of vertebrate striated muscles. By providing connections at the level of individual microfilaments, it contributes to the fine balance of forces between the two halves of the sarcomere. The Ig superfamily (IgSF) is a heterogenous group of proteins, built on a common fold comprised of a sandwich of two beta sheets. IgSF domains can be divided into 4 main classes based on their structures and sequences: the Variable (V), Constant 1 (C1), Constant 2 (C2), and Intermediate (I) sets. Unlike the V-set, one of the distinctive features of I-set domains is the lack of a C" strand. The structure of the titin Z1z2 lacks this strand and thus it belongs to the I-set of the IgSF. I-set domains are found in several cell adhesion molecules (such as VCAM, ICAM, and MADCAM), and are also present in numerous other diverse protein families, including several tyrosine-protein kinase receptors, the hemolymph protein hemolin, the muscle proteins titin, telokin, and twitchin, the neuronal adhesion molecule axonin-1, and the signaling molecule semaphorin 4D that is involved in axonal guidance, immune function and angiogenesis. Pssm-ID: 409564 [Multi-domain] Cd Length: 91 Bit Score: 53.74 E-value: 7.75e-09
|
|||||||
| TSP_1 | pfam00090 | Thrombospondin type 1 domain; |
254-301 | 1.04e-08 | |||
Thrombospondin type 1 domain; Pssm-ID: 459668 [Multi-domain] Cd Length: 49 Bit Score: 52.04 E-value: 1.04e-08
|
|||||||
| DEATH | smart00005 | DEATH domain, found in proteins involved in cell death (apoptosis); Alpha-helical domain ... |
855-927 | 1.96e-08 | |||
DEATH domain, found in proteins involved in cell death (apoptosis); Alpha-helical domain present in a variety of proteins with apoptotic functions. Some (but not all) of these domains form homotypic and heterotypic dimers. Pssm-ID: 214467 [Multi-domain] Cd Length: 88 Bit Score: 52.41 E-value: 1.96e-08
|
|||||||
| Ig5_Contactin | cd04969 | Fifth immunoglobulin (Ig) domain of contactin; The members here are composed of the fifth ... |
158-247 | 3.46e-08 | |||
Fifth immunoglobulin (Ig) domain of contactin; The members here are composed of the fifth immunoglobulin (Ig) domain of contactins. Contactins are neural cell adhesion molecules and are comprised of six Ig domains followed by four fibronectin type III (FnIII) domains anchored to the membrane by glycosylphosphatidylinositol. The first four Ig domains form the intermolecular binding fragment, which arranges as a compact U-shaped module via contacts between Ig domains 1 and 4, and between Ig domains 2 and 3. Contactin-2 (TAG-1, axonin-1) may play a part in the neuronal processes of neurite outgrowth, axon guidance and fasciculation, and neuronal migration. This group also includes contactin-1 and contactin-5. The different contactins show different expression patterns in the central nervous system. During development and in adulthood, contactin-2 is transiently expressed in subsets of central and peripheral neurons. Contactin-5 is expressed specifically in the rat postnatal nervous system, peaking at about 3 weeks postnatal, and a lack of contactin-5 (NB-2) results in an impairment of neuronal activity in the rat auditory system. Contactin-5 is highly expressed in the adult human brain in the occipital lobe and in the amygdala. Contactin-1 is differentially expressed in tumor tissues and may, through a RhoA mechanism, facilitate invasion and metastasis of human lung adenocarcinoma. Pssm-ID: 409358 [Multi-domain] Cd Length: 89 Bit Score: 51.69 E-value: 3.46e-08
|
|||||||
| IG_like | smart00410 | Immunoglobulin like; IG domains that cannot be classified into one of IGv1, IGc1, IGc2, IG. |
163-249 | 3.50e-08 | |||
Immunoglobulin like; IG domains that cannot be classified into one of IGv1, IGc1, IGc2, IG. Pssm-ID: 214653 [Multi-domain] Cd Length: 85 Bit Score: 51.74 E-value: 3.50e-08
|
|||||||
| TSP1_spondin | pfam19028 | Spondin-like TSP1 domain; This entry represents a sub-type of TSP1 domains that have an ... |
254-289 | 1.07e-07 | |||
Spondin-like TSP1 domain; This entry represents a sub-type of TSP1 domains that have an alternative disulphide binding pattern compared to the canonical TSP1 domain. Pssm-ID: 465948 Cd Length: 52 Bit Score: 49.20 E-value: 1.07e-07
|
|||||||
| Ig_Titin_like | cd05748 | Immunoglobulin (Ig)-like domain of titin and similar proteins; The members here are composed ... |
183-249 | 1.24e-07 | |||
Immunoglobulin (Ig)-like domain of titin and similar proteins; The members here are composed of the immunoglobulin (Ig)-like domain found in titin-like proteins and similar proteins. Titin (also called connectin) is a fibrous sarcomeric protein specifically found in vertebrate striated muscle. Titin is a giant protein; depending on isoform composition, it ranges from 2970 to 3700 kDa, and is of a length that spans half a sarcomere. Titin largely consists of multiple repeats of Ig-like and fibronectin type 3 (FN-III)-like domains. Titin connects the ends of myosin thick filaments to Z disks and extends along the thick filament to the H zone. It appears to function similarly to an elastic band, keeping the myosin filaments centered in the sarcomere during muscle contraction or stretching. Within the sarcomere, titin is also attached to or is associated with myosin binding protein C (MyBP-C). MyBP-C appears to contribute to the generation of passive tension by titin and like titin has repeated Ig-like and FN-III domains. Also included in this group are worm twitchin and insect projectin, thick filament proteins of invertebrate muscle which also have repeated Ig-like and FN-III domains. Pssm-ID: 409406 [Multi-domain] Cd Length: 82 Bit Score: 49.90 E-value: 1.24e-07
|
|||||||
| IgI_titin_I1-like | cd20951 | Immunoglobulin domain I1 of the titin I-band and similar proteins; a member of the I-set of ... |
171-247 | 1.47e-07 | |||
Immunoglobulin domain I1 of the titin I-band and similar proteins; a member of the I-set of IgSF domains; The members here are composed of the immunoglobulin domain I1 of the titin I-band and similar proteins. Titin is a key component in the assembly and functioning of vertebrate striated muscles. By providing connections at the level of individual microfilaments, it contributes to the fine balance of forces between the two halves of the sarcomere. The Ig superfamily (IgSF) is a heterogenous group of proteins, built on a common fold comprised of a sandwich of two beta sheets. The two sheets are linked together by a conserved disulfide bond between B strand and F strand. IgSF domains can be divided into 4 main classes based on their structures and sequences: the Variable (V), Constant 1 (C1), Constant 2 (C2), and Intermediate (I) sets. The Ig I1 domain of the titin I-band is a member of the I-set Ig domains, having A-B-E-D strands in one beta-sheet and A'-G-F-C-C' in the other. Like the V-set Ig domains, members of the I-set have a discontinuous A strand but lack a C" strand. I-set domains are found in several cell adhesion molecules (such as VCAM, ICAM, and MADCAM), and are also present in numerous other diverse protein families, including several tyrosine-protein kinase receptors, the hemolymph protein hemolin, the muscle proteins titin, telokin, and twitchin, the neuronal adhesion molecule axonin-1, and the signaling molecule semaphorin 4D that is involved in axonal guidance, immune function and angiogenesis. Pssm-ID: 409543 [Multi-domain] Cd Length: 94 Bit Score: 50.11 E-value: 1.47e-07
|
|||||||
| ig | pfam00047 | Immunoglobulin domain; Members of the immunoglobulin superfamily are found in hundreds of ... |
161-233 | 3.22e-07 | |||
Immunoglobulin domain; Members of the immunoglobulin superfamily are found in hundreds of proteins of different functions. Examples include antibodies, the giant muscle kinase titin and receptor tyrosine kinases. Immunoglobulin-like domains may be involved in protein-protein and protein-ligand interactions. Pssm-ID: 395002 Cd Length: 86 Bit Score: 49.12 E-value: 3.22e-07
|
|||||||
| Death_MyD88 | cd08312 | Death domain of Myeloid Differentation primary response protein MyD88; Death Domain (DD) of ... |
856-925 | 4.55e-07 | |||
Death domain of Myeloid Differentation primary response protein MyD88; Death Domain (DD) of Myeloid Differentiation primary response protein 88 (MyD88). MyD88 is an adaptor protein involved in interleukin-1 receptor (IL-1R)- and Toll-like receptor (TLR)-induced activation of nuclear factor-kappaB (NF-kB) and mitogen activated protein kinase pathways that lead to the induction of proinflammatory cytokines. It is a key component in the signaling pathway of pathogen recognition in the innate immune system. MyD88 contains an N-terminal DD and a C-terminal Toll/IL-1 Receptor (TIR) homology domain that mediates interaction with TLRs and IL-1R. In general, DDs are protein-protein interaction domains found in a variety of domain architectures. Their common feature is that they form homodimers by self-association or heterodimers by associating with other members of the DD superfamily including CARD (Caspase activation and recruitment domain), DED (Death Effector Domain), and PYRIN. They serve as adaptors in signaling pathways and can recruit other proteins into signaling complexes. Pssm-ID: 260026 Cd Length: 79 Bit Score: 48.37 E-value: 4.55e-07
|
|||||||
| IgI_3_NCAM-1 | cd05730 | Third immunoglobulin (Ig)-like domain of Neural Cell Adhesion Molecule 1 (NCAM-1); member of ... |
174-233 | 1.16e-06 | |||
Third immunoglobulin (Ig)-like domain of Neural Cell Adhesion Molecule 1 (NCAM-1); member of the I-set of IgSF domains; The members here are composed of the third immunoglobulin (Ig)-like domain of Neural Cell Adhesion Molecule (NCAM-1). NCAM plays important roles in the development and regeneration of the central nervous system, in synaptogenesis and neural migration. NCAM mediates cell-cell and cell-substratum recognition and adhesion via homophilic (NCAM-NCAM), and heterophilic (NCAM-non-NCAM), interactions. NCAM is expressed as three major isoforms having different intracellular extensions. The extracellular portion of NCAM has five N-terminal Ig-like domains and two fibronectin type III domains. The double zipper adhesion complex model for NCAM homophilic binding involves Ig1, Ig2, and Ig3. By this model, Ig1 and Ig2 mediate dimerization of NCAM molecules situated on the same cell surface (cis interactions), and Ig3 domains mediate interactions between NCAM molecules expressed on the surface of opposing cells (trans interactions) through binding to the Ig1 and Ig2 domains. The adhesive ability of NCAM is modulated by the addition of polysialic acid chains to the fifth Ig-like domain. Pssm-ID: 143207 [Multi-domain] Cd Length: 95 Bit Score: 47.62 E-value: 1.16e-06
|
|||||||
| IgI_3_Robo | cd05725 | Third immunoglobulin (Ig)-like domain in Robo (roundabout) receptors; member of the I-set of ... |
163-247 | 2.64e-06 | |||
Third immunoglobulin (Ig)-like domain in Robo (roundabout) receptors; member of the I-set of Ig superfamily (IgSF) domains; The members here are composed of the third immunoglobulin (Ig)-like domain in Robo (roundabout) receptors. Robo receptors play a role in the development of the central nervous system (CNS), and are receptors of Slit protein. Slit is a repellant secreted by the neural cells in the midline. Slit acts through Robo to prevent most neurons from crossing the midline from either side. Three mammalian Robo homologs (Robo1, Robo2, Robo3), and three mammalian Slit homologs (Slit-1,Slit-2, Slit-3), have been identified. Commissural axons, which cross the midline, express low levels of Robo; longitudinal axons, which avoid the midline, express high levels of Robo. Robo1, Robo2, and Robo3 are expressed by commissural neurons in the vertebrate spinal cord and Slit-1, Slit-2, and Slit-3 are expressed at the ventral midline. Robo-3 is a divergent member of the Robo family which instead of being a positive regulator of Slit responsiveness, antagonizes Slit responsiveness in precrossing axons. The Slit-Robo interaction is mediated by the second leucine-rich repeat (LRR) domain of Slit and the two N-terminal Ig domains of Robo, Ig1 and Ig2. The primary Robo binding site for Slit2 has been shown by surface plasmon resonance experiments and mutational analysis to be the Ig1 domain, while the Ig2 domain has been proposed to harbor a weak secondary binding site. This group belongs to the I-set of IgSF domains, having A-B-E-D strands in one beta-sheet and A'-G-F-C-C' in the other. Like the V-set Ig domains, members of the I-set have a discontinuous A strand, but lack a C" strand. Pssm-ID: 409390 [Multi-domain] Cd Length: 83 Bit Score: 46.23 E-value: 2.64e-06
|
|||||||
| Ig6_Contactin | cd04970 | Sixth immunoglobulin (Ig) domain of contactin; The members here are composed of the sixth ... |
191-247 | 3.63e-06 | |||
Sixth immunoglobulin (Ig) domain of contactin; The members here are composed of the sixth immunoglobulin (Ig) domain of contactins. Contactins are neural cell adhesion molecules and are comprised of six Ig domains followed by four fibronectin type III (FnIII) domains anchored to the membrane by glycosylphosphatidylinositol. The first four Ig domains form the intermolecular binding fragment, which arranges as a compact U-shaped module via contacts between Ig domains 1 and 4, and between Ig domains 2 and 3. Contactin-2 (TAG-1, axonin-1) may play a part in the neuronal processes of neurite outgrowth, axon guidance and fasciculation, and neuronal migration. This group also includes contactin-1 and contactin-5. The different contactins show different expression patterns in the central nervous system. During development and in adulthood, contactin-2 is transiently expressed in subsets of central and peripheral neurons. Contactin-5 is expressed specifically in the rat postnatal nervous system, peaking at about 3 weeks postnatal, and a lack of contactin-5 (NB-2) results in an impairment of neuronal activity in the rat auditory system. Contactin-5 is highly expressed in the adult human brain in the occipital lobe and in the amygdala. Contactin-1 is differentially expressed in tumor tissues and may, through a RhoA mechanism, facilitate invasion and metastasis of human lung adenocarcinoma. Pssm-ID: 409359 Cd Length: 102 Bit Score: 46.39 E-value: 3.63e-06
|
|||||||
| Death_DAPK1 | cd08782 | Death domain found in death-associated protein kinase 1; Death domain (DD) found in ... |
855-925 | 4.76e-06 | |||
Death domain found in death-associated protein kinase 1; Death domain (DD) found in death-associated protein kinase 1 (DAPK1). DAPK1 is composed of several functional domains, including a kinase domain, a CaM regulatory domain, ankyrin repeats, a cytoskeletal-binding domain and a C-terminal DD. It plays important roles in a diverse range of signal transduction pathways including apoptosis, growth factor signalling, and autophagy. Loss of DAPK1 expression, usually because of DNA methylation, is implicated in many tumor types. DAPK1 is highly abundant in the brain and has also been associated with neurodegeneration. In general, DDs are protein-protein interaction domains found in a variety of domain architectures. Their common feature is that they form homodimers by self-association or heterodimers by associating with other members of the DD superfamily including CARD (Caspase activation and recruitment domain), DED (Death Effector Domain), and PYRIN. They serve as adaptors in signaling pathways and can recruit other proteins into signaling complexes. Pssm-ID: 260052 Cd Length: 82 Bit Score: 45.41 E-value: 4.76e-06
|
|||||||
| IgI_VEGFR | cd04976 | Immunoglobulin (Ig)-like domain of vascular endothelial growth factor receptor (VEGFR); member ... |
183-247 | 5.02e-06 | |||
Immunoglobulin (Ig)-like domain of vascular endothelial growth factor receptor (VEGFR); member of the I-set of Ig superfamily (IgSF) domains; The members here are composed of the immunoglobulin (Ig)-like domain of vascular endothelial growth factor receptor (VEGFR). The VEGFRs have an extracellular component with seven Ig-like domains, a transmembrane segment, and an intracellular tyrosine kinase domain interrupted by a kinase-insert domain. The VEGFR family consists of three members, VEGFR-1 (Flt-1), VEGFR-2 (KDR/Flk-1), and VEGFR-3 (Flt-4). VEGFRs bind VEGFs with high affinity at the Ig-like domains. VEGF-A is important to the growth and maintenance of vascular endothelial cells and to the development of new blood- and lymphatic-vessels in physiological and pathological states. VEGFR-2 is a major mediator of the mitogenic, angiogenic, and microvascular permeability-enhancing effects of VEGF-A. VEGFR-1 may play an inhibitory part in these processes by binding VEGF and interfering with its interaction with VEGFR-2. VEGFR-1 has a signaling role in mediating monocyte chemotaxis. VEGFR-1 and VEGFR-2 may mediate a chemotactic and a survival signal in hematopoietic stem cells or leukemia cells. VEGFR-3 has been shown to be involved in tumor angiogenesis and growth. This group belongs to the I-set of IgSF domains, having A-B-E-D strands in one beta-sheet and A'-G-F-C-C' in the other. Like the V-set Ig domains, members of the I-set have a discontinuous A strand, but lack a C" strand. Pssm-ID: 409365 Cd Length: 90 Bit Score: 45.67 E-value: 5.02e-06
|
|||||||
| IgI_Myotilin_C_like | cd05744 | Immunoglobulin (Ig)-like domain of myotilin, palladin, and myopalladin; member of the I-set of ... |
159-233 | 6.10e-06 | |||
Immunoglobulin (Ig)-like domain of myotilin, palladin, and myopalladin; member of the I-set of Ig superfamily (IgSF) domains; The members here are composed of the immunoglobulin (Ig)-like domain in myotilin, palladin, and myopalladin. Myotilin, palladin, and myopalladin function as scaffolds that regulate actin organization. Myotilin and myopalladin are most abundant in skeletal and cardiac muscle; palladin is ubiquitously expressed in the organs of developing vertebrates and plays a key role in cellular morphogenesis. The three family members each interact with specific molecular partners with all three binding to alpha-actinin; In addition, palladin also binds to vasodilator-stimulated phosphoprotein (VASP) and ezrin, myotilin binds to filamin and actin, and myopalladin also binds to nebulin and cardiac ankyrin repeat protein (CARP). This group belongs to the I-set of IgSF domains, having A-B-E-D strands in one beta-sheet and A'-G-F-C-C' in the other. Like the V-set Ig domains, members of the I-set have a discontinuous A strand, but lack a C" strand. Pssm-ID: 409405 [Multi-domain] Cd Length: 91 Bit Score: 45.56 E-value: 6.10e-06
|
|||||||
| IgI_4_hemolin-like | cd20978 | Fourth immunoglobulin (Ig)-like domain of hemolin, and similar domains; a member of the I-set ... |
174-247 | 6.90e-06 | |||
Fourth immunoglobulin (Ig)-like domain of hemolin, and similar domains; a member of the I-set of IgSF domains; The members here are composed of the fourth immunoglobulin (Ig)-like domain of hemolin and similar proteins. Hemolin, an insect immunoglobulin superfamily (IgSF) member containing four Ig-like domains, is a lipopolysaccharide-binding immune protein induced during bacterial infection. Hemolin shares significant sequence similarity with the first four Ig-like domains of the transmembrane cell adhesion molecules (CAMs) of the L1 family. IgSF domains can be divided into 4 main classes based on their structures and sequences: the Variable (V), Constant 1 (C1), Constant 2 (C2), and Intermediate (I) sets. The fourth Ig-like domain of hemolin is a member of the I-set Ig domains, having A-B-E-D strands in one beta-sheet and A'-G-F-C-C' in the other. Like the V-set domains, members of the I-set have a discontinuous A strand but lack a C" strand. I-set domains are found in several cell adhesion molecules (such as VCAM, ICAM, and MADCAM), and are also present in numerous other diverse protein families, including several tyrosine-protein kinase receptors, the muscle proteins titin, telokin, and twitchin, the neuronal adhesion molecule axonin-1, and the signaling molecule semaphorin 4D that is involved in axonal guidance, immune function and angiogenesis. Pssm-ID: 409570 [Multi-domain] Cd Length: 88 Bit Score: 45.08 E-value: 6.90e-06
|
|||||||
| Ig_2 | pfam13895 | Immunoglobulin domain; This domain contains immunoglobulin-like domains. |
168-247 | 1.61e-05 | |||
Immunoglobulin domain; This domain contains immunoglobulin-like domains. Pssm-ID: 464026 [Multi-domain] Cd Length: 79 Bit Score: 43.92 E-value: 1.61e-05
|
|||||||
| IgI_1_MuSK | cd20970 | agrin-responsive first immunoglobulin-like domains (Ig1) of the MuSK ectodomain; a member of ... |
170-247 | 3.92e-05 | |||
agrin-responsive first immunoglobulin-like domains (Ig1) of the MuSK ectodomain; a member of the I-set of IgSF domains; The members here are composed of the first immunoglobulin-like domains (Ig1) of the Muscle-specific kinase (MuSK). MuSK is a receptor tyrosine kinase specifically expressed in skeletal muscle, where it plays a central role in the formation and maintenance of the neuromuscular junction (NMJ). MuSK is activated by agrin, a neuron-derived heparan sulfate proteoglycan. The activation of MUSK in myotubes regulates the formation of NMJs through the regulation of different processes including the specific expression of genes in subsynaptic nuclei, the reorganization of the actin cytoskeleton and the clustering of the acetylcholine receptors (AChR) in the postsynaptic membrane. The Ig superfamily (IgSF) is a heterogenous group of proteins, built on a common fold comprised of a sandwich of two beta sheets. IgSF domains can be divided into 4 main classes based on their structures and sequences: the Variable (V), Constant 1 (C1), Constant 2 (C2), and Intermediate (I) sets. Unlike the V-set, one of the distinctive features of I-set domains is the lack of a C" strand. The structure of the MuSK lacks this strand and thus it belongs to the I-set of the IgSF. I-set domains are found in several cell adhesion molecules (such as VCAM, ICAM, and MADCAM), and are also present in numerous other diverse protein families, including several tyrosine-protein kinase receptors, the hemolymph protein hemolin, the muscle proteins titin, telokin, and twitchin, the neuronal adhesion molecule axonin-1, and the signaling molecule semaphorin 4D that is involved in axonal guidance, immune function and angiogenesis. Pssm-ID: 409562 [Multi-domain] Cd Length: 92 Bit Score: 43.27 E-value: 3.92e-05
|
|||||||
| IgI_1_Contactin-1 | cd05849 | First immunoglobulin (Ig) domain of contactin-1; member of the I-set of Ig superfamily domains; ... |
159-245 | 4.06e-05 | |||
First immunoglobulin (Ig) domain of contactin-1; member of the I-set of Ig superfamily domains; The members here are composed of the first immunoglobulin (Ig) domain of the neural cell adhesion molecule contactin-1. Contactins are comprised of six Ig domains followed by four fibronectin type III (FnIII) domains anchored to the membrane by glycosylphosphatidylinositol. Contactin-1 is differentially expressed in tumor tissues and may, through a RhoA mechanism, facilitate invasion and metastasis of human lung adenocarcinoma. This group belongs to the I-set of IgSF domains. Pssm-ID: 409436 [Multi-domain] Cd Length: 95 Bit Score: 43.40 E-value: 4.06e-05
|
|||||||
| IgI_1_Contactin | cd04967 | First immunoglobulin (Ig) domain of contactin; member of the I-set of (Ig) superfamily domains; ... |
159-245 | 4.38e-05 | |||
First immunoglobulin (Ig) domain of contactin; member of the I-set of (Ig) superfamily domains; The members here are composed of the first immunoglobulin (Ig) domain of contactins. Contactins are neural cell adhesion molecules and are comprised of six Ig domains followed by four fibronectin type III (FnIII) domains anchored to the membrane by glycosylphosphatidylinositol. The first four Ig domains form the intermolecular binding fragment, which arranges as a compact U-shaped module via contacts between Ig domains 1 and 4, and between Ig domains 2 and 3. Contactin-2 (TAG-1, axonin-1) may play a part in the neuronal processes of neurite outgrowth, axon guidance and fasciculation, and neuronal migration. This group also includes contactin-1 and contactin-5. The different contactins show different expression patterns in the central nervous system. During development and in adulthood, contactin-2 is transiently expressed in subsets of central and peripheral neurons. Contactin-5 is expressed specifically in the rat postnatal nervous system, peaking at about 3 weeks postnatal, and a lack of contactin-5 (NB-2) results in an impairment of neuronal activity in the rat auditory system. Contactin-5 is highly expressed in the adult human brain in the occipital lobe and in the amygdala. Contactin-1 is differentially expressed in tumor tissues and may, through a RhoA mechanism, facilitate invasion and metastasis of human lung adenocarcinoma. This group belongs to the I-set of IgSF domains. Pssm-ID: 409356 [Multi-domain] Cd Length: 96 Bit Score: 43.00 E-value: 4.38e-05
|
|||||||
| TSP_1 | pfam00090 | Thrombospondin type 1 domain; |
311-357 | 5.45e-05 | |||
Thrombospondin type 1 domain; Pssm-ID: 459668 [Multi-domain] Cd Length: 49 Bit Score: 41.25 E-value: 5.45e-05
|
|||||||
| Death_NFkB-like | cd08310 | Death domain of Nuclear Factor-KappaB precursor proteins; Death Domain (DD) of Nuclear ... |
855-926 | 5.57e-05 | |||
Death domain of Nuclear Factor-KappaB precursor proteins; Death Domain (DD) of Nuclear Factor-KappaB (NF-kB) precursor proteins. The NF-kB family of transcription factors play a central role in cardiovascular growth, stress response, and inflammation by controlling the expression of a network of different genes. There are five NF-kB proteins, all containing an N-terminal REL Homology Domain (RHD). Two of these, NF-kB1 and NF-kB2 are produced from the processing of the precursor proteins p105 and p100, respectively. In addition to RHD, p105 and p100 contain ANK repeats and a C-terminal DD. NF-kBs are regulated by the Inhibitor of NF-kB (IkB) Kinase (IKK) complex through classical and non-canonical pathways, which differ in the IKK subunits involved and downstream targets. IKKs facilitate the release of NF-kB dimers from an inactive state, allowing them to migrate to the nucleus where they regulate gene transcription. The precursor proteins p105 and p100 function as IkBs and as NF-kB proteins after being processed by the proteasome. In general, DDs are protein-protein interaction domains found in a variety of domain architectures. Their common feature is that they form homodimers by self-association or heterodimers by associating with other members of the DD superfamily including CARD (Caspase activation and recruitment domain), DED (Death Effector Domain), and PYRIN. They serve as adaptors in signaling pathways and can recruit other proteins into signaling complexes. Pssm-ID: 260024 Cd Length: 72 Bit Score: 42.23 E-value: 5.57e-05
|
|||||||
| Ig2_Contactin-2-like | cd05727 | Second Ig domain of the neural cell adhesion molecule contactin-2, and similar domains; The ... |
174-235 | 1.08e-04 | |||
Second Ig domain of the neural cell adhesion molecule contactin-2, and similar domains; The members here are composed of the second Ig domain of the neural cell adhesion molecule contactin-2. Contactins are comprised of six Ig domains followed by four fibronectin type III (FnIII) domains anchored to the membrane by glycosylphosphatidylinositol. Contactin-2 (also called TAG-1, axonin-1) facilitates cell adhesion by homophilic binding between molecules in apposed membranes. The first four Ig domains form the intermolecular binding fragment which arranges as a compact U-shaped module by contacts between Ig domains 1 and 4, and domains 2 and 3. It has been proposed that a linear zipper-like array forms, from contactin-2 molecules alternatively provided by the two apposed membranes. Pssm-ID: 409392 Cd Length: 88 Bit Score: 41.78 E-value: 1.08e-04
|
|||||||
| IgI_telokin-like | cd20973 | immunoglobulin-like domain of telokin and similar proteins; a member of the I-set of IgSF ... |
182-247 | 1.40e-04 | |||
immunoglobulin-like domain of telokin and similar proteins; a member of the I-set of IgSF domains; The members here are composed of the immunoglobulin (Ig) domain in telokin, the C-terminal domain of myosin light chain kinase which is identical to telokin, and similar proteins. The Ig superfamily (IgSF) is a heterogenous group of proteins, built on a common fold comprised of a sandwich of two beta sheets. IgSF domains can be divided into 4 main classes based on their structures and sequences: the Variable (V), Constant 1 (C1), Constant 2 (C2), and Intermediate (I) sets. Unlike the V-set, one of the distinctive features of I-set domains is the lack of a C" strand. The structure of the telokin Ig domain lacks this strand and thus it belongs to the I-set of the IgSF. I-set domains are found in several cell adhesion molecules (such as VCAM, ICAM, and MADCAM), and are also present in numerous other diverse protein families, including several tyrosine-protein kinase receptors, the hemolymph protein hemolin, the muscle proteins titin, telokin, and twitchin, the neuronal adhesion molecule axonin-1, and the signaling molecule semaphorin 4D that is involved in axonal guidance, immune function and angiogenesis. Pssm-ID: 409565 [Multi-domain] Cd Length: 88 Bit Score: 41.41 E-value: 1.40e-04
|
|||||||
| IgI_2_FGFR_like | cd05729 | Second immunoglobulin (Ig)-like domain of fibroblast growth factor (FGF) receptor, and similar ... |
168-233 | 1.47e-04 | |||
Second immunoglobulin (Ig)-like domain of fibroblast growth factor (FGF) receptor, and similar domains; member of the I-set of IgSF domains; The members here are composed of the second immunoglobulin (Ig)-like domain of fibroblast growth factor (FGF) receptor. FGF receptors bind FGF signaling polypeptides. FGFs participate in multiple processes such as morphogenesis, development, and angiogenesis. FGFs bind to four FGF receptor tyrosine kinases (FGFR1, FGFR2, FGFR3, FGFR4). Receptor diversity is controlled by alternative splicing producing splice variants with different ligand binding characteristics and different expression patterns. FGFRs have an extracellular region comprised of three Ig-like domains, a single transmembrane helix, and an intracellular tyrosine kinase domain. Ligand binding and specificity reside in the Ig-like domains 2 and 3, and the linker region that connects these two. FGFR activation and signaling depend on FGF-induced dimerization, a process involving cell surface heparin or heparin sulfate proteoglycans. This group also contains fibroblast growth factor (FGF) receptor like-1(FGFRL1). FGFRL1 does not have a protein tyrosine kinase domain at its C-terminus; neither does its cytoplasmic domain appear to interact with a signaling partner. It has been suggested that FGFRL1 may not have any direct signaling function, but instead acts as a decoy receptor trapping FGFs and preventing them from binding other receptors. Pssm-ID: 409393 [Multi-domain] Cd Length: 95 Bit Score: 41.82 E-value: 1.47e-04
|
|||||||
| IgI_1_Contactin-5 | cd05848 | First immunoglobulin (Ig) domain of contactin-5; member of the I-set of Ig superfamily domains; ... |
159-245 | 1.57e-04 | |||
First immunoglobulin (Ig) domain of contactin-5; member of the I-set of Ig superfamily domains; The members here are composed of the first immunoglobulin (Ig) domain of the neural cell adhesion molecule contactin-5. Contactins are comprised of six Ig domains followed by four fibronectin type III (FnIII) domains, anchored to the membrane by glycosylphosphatidylinositol. The different contactins show different expression patterns in the central nervous system. In rats, a lack of contactin-5 (NB-2) results in an impairment of the neuronal activity in the auditory system. Contactin-5 is expressed specifically in the postnatal nervous system, peaking at about 3 weeks postnatal. Contactin-5 is highly expressed in the adult human brain in the occipital lobe and in the amygdala; lower levels of expression have been detected in the corpus callosum, caudate nucleus, and spinal cord. This group belongs to the I-set of IgSF domains. Pssm-ID: 409435 Cd Length: 96 Bit Score: 41.47 E-value: 1.57e-04
|
|||||||
| TSP1_ADAMTS | pfam19030 | Thrombospondin type 1 domain; This subfamily of thrombospondin type 1 repeats are mainly found ... |
257-290 | 1.57e-04 | |||
Thrombospondin type 1 domain; This subfamily of thrombospondin type 1 repeats are mainly found in ADAMTS proteins. Pssm-ID: 465950 [Multi-domain] Cd Length: 55 Bit Score: 40.51 E-value: 1.57e-04
|
|||||||
| Ig_Sema3 | cd05871 | Immunoglobulin (Ig)-like domain of class III semaphorin Sema3; The members here are composed ... |
176-231 | 1.88e-04 | |||
Immunoglobulin (Ig)-like domain of class III semaphorin Sema3; The members here are composed of the immunoglobulin (Ig)-like domain of Sema3 and similar proteins. Semaphorins are classified based on structural features additional to the Sema domain. Sema3 is a Class III semaphorin that is secreted. It is a vertebrate class having a Sema domain, an Ig domain, a short basic domain. They have been shown to be axonal guidance cues and have a part in the regulation of the cardiovascular, immune, and respiratory systems. Sema3A, the prototype member of this class III subfamily, induces growth cone collapse and is an inhibitor of axonal sprouting. In perinatal rat cortex, it acts as a chemoattractant and functions to direct the orientated extension of apical dendrites. It may play a role, prior to the development of apical dendrites, in signaling the radial migration of newborn cortical neurons towards the upper layers. Sema3A selectively inhibits vascular endothelial growth factor receptor (VEGF)-induced angiogenesis and induces microvascular permeability. This group also includes Sema3B, -C, -D, -E, -G. Pssm-ID: 409455 Cd Length: 92 Bit Score: 41.18 E-value: 1.88e-04
|
|||||||
| Ig_Pro_neuregulin-1 | cd05895 | Immunoglobulin (Ig)-like domain found in neuregulin (NRG)-1; The members here are composed of ... |
166-247 | 1.93e-04 | |||
Immunoglobulin (Ig)-like domain found in neuregulin (NRG)-1; The members here are composed of the immunoglobulin (Ig)-like domain found in neuregulin (NRG)-1. There are many NRG-1 isoforms which arise from the alternative splicing of mRNA. NRG-1 belongs to the neuregulin gene family which is comprised of four genes. This group represents NRG-1. NRGs are signaling molecules which participate in cell-cell interactions in the nervous system, breast, and heart, and other organ systems, and are implicated in the pathology of diseases including schizophrenia, multiple sclerosis, and breast cancer. The NRG-1 protein binds to and activates the tyrosine kinases receptors ErbB3 and ErbB4, initiating signaling cascades. NRG-1 has multiple functions, for example, in the brain it regulates various processes such as radial glia formation and neuronal migration, dendritic development, and expression of neurotransmitters receptors in the peripheral nervous system NRG-1 regulates processes such as target cell differentiation, and Schwann cell survival. Pssm-ID: 409476 Cd Length: 93 Bit Score: 41.13 E-value: 1.93e-04
|
|||||||
| IgI_2_L1-CAM_like | cd05845 | Second immunoglobulin (Ig)-like domain of the L1 cell adhesion molecule (CAM), and similar ... |
174-232 | 2.12e-04 | |||
Second immunoglobulin (Ig)-like domain of the L1 cell adhesion molecule (CAM), and similar domains; member of the I-set of Ig superfamily (IgSF) domains; The members here are composed of the second immunoglobulin (Ig)-like domain of the L1 cell adhesion molecule (CAM) and similar proteins. L1 belongs to the L1 subfamily of cell adhesion molecules (CAMs) and is comprised of an extracellular region having six Ig-like domains, five fibronectin type III domains, a transmembrane region, and an intracellular domain. L1 is primarily expressed in the nervous system and is involved in its development and function. L1 is associated with an X-linked recessive disorder, X-linked hydrocephalus, MASA syndrome, or spastic paraplegia type 1 that involves abnormalities of axonal growth. This group belongs to the I-set of IgSF domains, having A-B-E-D strands in one beta-sheet and A'-G-F-C-C' in the other. Like the V-set Ig domains, members of the I-set have a discontinuous A strand, but lack a C" strand. Pssm-ID: 409432 Cd Length: 91 Bit Score: 40.94 E-value: 2.12e-04
|
|||||||
| IgI_2_RPTP_IIa_LAR_like | cd05738 | Second immunoglobulin (Ig)-like domain of the receptor protein tyrosine phosphatase (RPTP)-F; ... |
183-247 | 2.14e-04 | |||
Second immunoglobulin (Ig)-like domain of the receptor protein tyrosine phosphatase (RPTP)-F; member of the I-set of IgSF domains; The members here are composed of the second immunoglobulin (Ig)-like domain found in the receptor protein tyrosine phosphatase (RPTP)-F, also known as LAR. LAR belongs to the RPTP type IIa subfamily. Members of this subfamily are cell adhesion molecule-like proteins involved in central nervous system (CNS) development. They have large extracellular portions comprised of multiple Ig-like domains and two to nine fibronectin type III (FNIII) domains and a cytoplasmic portion having two tandem phosphatase domains. Pssm-ID: 409400 [Multi-domain] Cd Length: 91 Bit Score: 41.15 E-value: 2.14e-04
|
|||||||
| Ig4_Contactin-2-like | cd05728 | Fourth Ig domain of the neural cell adhesion molecule contactin-2, and similar domains; The ... |
159-233 | 2.64e-04 | |||
Fourth Ig domain of the neural cell adhesion molecule contactin-2, and similar domains; The members here are composed of the fourth Ig domain of the neural cell adhesion molecule contactin-2. Contactins are comprised of six Ig domains followed by four fibronectin type III (FnIII) domains anchored to the membrane by glycosylphosphatidylinositol. Contactin-2 (also called TAG-1, axonin-1) facilitates cell adhesion by homophilic binding between molecules in apposed membranes. The first four Ig domains form the intermolecular binding fragment which arranges as a compact U-shaped module by contacts between Ig domains 1 and 4, and domains 2 and 3. It has been proposed that a linear zipper-like array forms, from contactin-2 molecules alternatively provided by the two apposed membranes. Pssm-ID: 143205 [Multi-domain] Cd Length: 85 Bit Score: 40.66 E-value: 2.64e-04
|
|||||||
| IgI_VEGFR-2 | cd05864 | Immunoglobulin (Ig)-like domain of vascular endothelial growth factor receptor 2 (VEGFR-2); ... |
183-245 | 2.65e-04 | |||
Immunoglobulin (Ig)-like domain of vascular endothelial growth factor receptor 2 (VEGFR-2); member of the I-set of Ig superfamily (IgSF) domains; The members here are composed of the immunoglobulin (Ig)-like domain of vascular endothelial growth factor receptor 2 (VEGFR-2). The VEGFRs have an extracellular component with seven Ig-like domains, a transmembrane segment, and an intracellular tyrosine kinase domain interrupted by a kinase-insert domain. VEGFRs bind VEGFs with high affinity at the Ig-like domains. VEGFR-2 (KDR/Flk-1) is a major mediator of the mitogenic, angiogenic and microvascular permeability-enhancing effects of VEGF-A; VEGF-A is important to the growth and maintenance of vascular endothelial cells and to the development of new blood- and lymphatic-vessels in physiological and pathological states. VEGF-A also interacts with VEGFR-1, which it binds more strongly than VEGFR-2. VEGFR-1 and VEGFR-2 may mediate a chemotactic and a survival signal in hematopoietic stem cells or leukemia cells. This group belongs to the I-set of IgSF domains, having A-B-E-D strands in one beta-sheet and A'-G-F-C-C' in the other. Like the V-set Ig domains, members of the I-set have a discontinuous A strand, but lack a C" strand. Pssm-ID: 409450 Cd Length: 89 Bit Score: 40.68 E-value: 2.65e-04
|
|||||||
| IgI_2_FGFR | cd05857 | Second immunoglobulin (Ig)-like domain of fibroblast growth factor (FGF) receptor; member of ... |
168-233 | 2.78e-04 | |||
Second immunoglobulin (Ig)-like domain of fibroblast growth factor (FGF) receptor; member of the I-set of IgSF domains; The members here are composed of the second immunoglobulin (Ig)-like domain of fibroblast growth factor (FGF) receptor. FGF receptors bind FGF signaling polypeptides. FGFs participate in multiple processes such as morphogenesis, development, and angiogenesis. FGFs bind to four FGF receptor tyrosine kinases (FGFR1, FGFR2, FGFR3, FGFR4). Receptor diversity is controlled by alternative splicing producing splice variants with different ligand binding characteristics and different expression patterns. FGFRs have an extracellular region comprised of three IG-like domains, a single transmembrane helix, and an intracellular tyrosine kinase domain. Ligand binding and specificity reside in the Ig-like domains 2 and 3, and the linker region that connects these two. FGFR activation and signaling depend on FGF-induced dimerization, a process involving cell surface heparin or heparin sulfate proteoglycans. Pssm-ID: 409443 [Multi-domain] Cd Length: 95 Bit Score: 40.99 E-value: 2.78e-04
|
|||||||
| IgC_1_Robo | cd07693 | First immunoglobulin (Ig)-like constant domain in Robo (roundabout) receptors, and similar ... |
176-247 | 2.89e-04 | |||
First immunoglobulin (Ig)-like constant domain in Robo (roundabout) receptors, and similar domains; The members here are composed of the first immunoglobulin (Ig)-like domain in Roundabout (Robo) receptors. Robo receptors play a role in the development of the central nervous system (CNS), and are receptors of Slit protein. Slit is a repellant secreted by the neural cells in the midline. Slit acts through Robo to prevent most neurons from crossing the midline from either side. Three mammalian Robo homologs (Robo1, Robo2, and Robo3), and three mammalian Slit homologs (Slit1, Slit2, Slit3), have been identified. Commissural axons, which cross the midline, express low levels of Robo; longitudinal axons, which avoid the midline, express high levels of Robo. Robo1, Robo2, and Robo3 are expressed by commissural neurons in the vertebrate spinal cord and Slit1, Slit2,and Slit3 are expressed at the ventral midline. Robo3 is a divergent member of the Robo family which instead of being a positive regulator of Slit responsiveness, antagonizes Slit responsiveness in precrossing axons. The Slit-Robo interaction is mediated by the second leucine-rich repeat (LRR) domain of Slit and the two N-terminal Ig domains of Robo, Ig1 and Ig2. The primary Robo binding site for Slit2 has been shown by surface plasmon resonance experiments and mutational analysis to be is the Ig1 domain, while the Ig2 domain has been proposed to harbor a weak secondary binding site. Pssm-ID: 409490 [Multi-domain] Cd Length: 99 Bit Score: 41.00 E-value: 2.89e-04
|
|||||||
| IgI_4_MYLK-like | cd20976 | Fourth Ig-like domain from smooth muscle myosin light chain kinase and similar domains ; a ... |
170-249 | 5.18e-04 | |||
Fourth Ig-like domain from smooth muscle myosin light chain kinase and similar domains ; a member of the I-set of IgSF domains; The members here are composed of the fourth immunoglobulin (Ig)-like domain from smooth muscle myosin light chain kinase (MYLK) and similar domains. The Ig superfamily (IgSF) is a heterogenous group of proteins, built on a common fold comprised of a sandwich of two beta sheets. IgSF domains can be divided into 4 main classes based on their structures and sequences: the Variable (V), Constant 1 (C1), Constant 2 (C2), and Intermediate (I) sets. Unlike the V-set, one of the distinctive features of I-set domains is the lack of a C" strand. The structure of this group shows that the fourth Ig-like domain from myosin light chain kinase lacks this strand and thus belongs to the I-set of the IgSF. I-set domains are found in several cell adhesion molecules (such as VCAM, ICAM, and MADCAM), and are also present in numerous other diverse protein families, including several tyrosine-protein kinase receptors, the hemolymph protein hemolin, the muscle proteins titin, telokin, and twitchin, the neuronal adhesion molecule axonin-1, and the signaling molecule semaphorin 4D that is involved in axonal guidance, immune function and angiogenesis. Pssm-ID: 409568 [Multi-domain] Cd Length: 90 Bit Score: 39.93 E-value: 5.18e-04
|
|||||||
| IgI_6_Dscam | cd20959 | Sixth immunoglobulin domain of the Drosophila melanogaster Dscam protein, and similar domains; ... |
173-247 | 6.04e-04 | |||
Sixth immunoglobulin domain of the Drosophila melanogaster Dscam protein, and similar domains; a member of the I-set of IgSF domains; The members here are composed of the sixth immunoglobulin domain of the Drosophila melanogaster Down syndrome cell adhesion molecule (DSCAM) protein and similar proteins. Down syndrome cell adhesion molecule (DSCAM) is a cell adhesion molecule that plays critical roles in neural development, including axon guidance and branching, axon target recognition, self-avoidance and synaptic formation. DSCAM belongs to the immunoglobulin superfamily and contributes to defects in the central nervous system in Down syndrome patients. Vertebrate DSCAMs differ from Drosophila Dscam1 in that they lack the extensive alternative splicing that occurs in the insect gene. Drosophila melanogaster Dscam has 38,016 isoforms generated by the alternative splicing of four variable exon clusters, which allows every neuron in the fly to display a distinctive set of Dscam proteins on its cell surface. Drosophila Dscam1 is a cell-surface protein that plays important roles in neural development and axon tiling of neurons. It is shown that thousands of isoforms bind themselves through specific homophilic (self-binding) interactions, a process which mediates cellular self-recognition. Drosophila Dscam2 is also alternatively spliced and plays a key role in the development of two visual system neurons, monopolar cells L1 and L2. This group is a member of the I-set Ig domains, having A-B-E-D strands in one beta-sheet and A'-G-F-C-C' in the other. Like the V-set Ig domains, members of the I-set have a discontinuous A strand but lack a C" strand. Pssm-ID: 409551 Cd Length: 94 Bit Score: 39.78 E-value: 6.04e-04
|
|||||||
| IgI_Lingo-1 | cd20969 | Immunoglobulin I-set domain of the Leucine-rich repeat and immunoglobin-like domain-containing ... |
174-233 | 6.29e-04 | |||
Immunoglobulin I-set domain of the Leucine-rich repeat and immunoglobin-like domain-containing protein 1 (Lingo-1); The members here are composed of the immunoglobulin I-set (IgI) domain of the Leucine-rich repeat and immunoglobin-like domain-containing protein 1 (Lingo-1). Human Lingo-1 is a central nervous system-specific transmembrane glycoprotein also known as LERN-1, which functions as a negative regulator of neuronal survival, axonal regeneration, and oligodendrocyte differentiation and myelination. Lingo-1 is a key component of the Nogo receptor signaling complex (RTN4R/NGFR) in RhoA activation responsible for some inhibition of axonal regeneration by myelin-associated factors. The ligand-binding ectodomain of human Lingo-1 contains a bimodular, kinked structure composed of leucine-rich repeat (LRR) and immunoglobulin (Ig)-like modules. Diseases associated with Lingo-1 include mental retardation, autosomal recessive 64 and essential tremor. The Ig superfamily (IgSF) is a heterogenous group of proteins, built on a common fold comprised of a sandwich of two beta sheets. IgSF domains can be divided into 4 main classes based on their structures and sequences: the Variable (V), Constant 1 (C1), Constant 2 (C2), and Intermediate (I) sets. Unlike the V-set, one of the distinctive features of I-set domains is the lack of a C" strand. The structure of the Lingo-1 lacks this strand and thus it belongs to the I-set of the IgSF. I-set domains are found in several cell adhesion molecules (such as VCAM, ICAM, and MADCAM), and are also present in numerous other diverse protein families, including several tyrosine-protein kinase receptors, the hemolymph protein hemolin, the muscle proteins titin, telokin, and twitchin, the neuronal adhesion molecule axonin-1, and the signaling molecule semaphorin 4D that is involved in axonal guidance, immune function and angiogenesis. Pssm-ID: 409561 Cd Length: 92 Bit Score: 39.68 E-value: 6.29e-04
|
|||||||
| IgI_2_JAM1 | cd20950 | Second Ig-like domain of Junctional adhesion molecule-1 (JAM1); a member of the I-set of IgSF ... |
168-233 | 1.67e-03 | |||
Second Ig-like domain of Junctional adhesion molecule-1 (JAM1); a member of the I-set of IgSF domains; The members here are composed of the second Ig-like domain of Junctional adhesion molecule-1 (JAM1). JAM1 is an immunoglobulin superfamily (IgSF) protein with two Ig-like domains in its extracellular region; it plays a role in the formation of endothelial and epithelial tight junction and acts as a receptor for mammalian reovirus sigma-1. The IgSF is a heterogenous group of proteins, built on a common fold comprised of a sandwich of two beta sheets. The two sheets are linked together by a conserved disulfide bond between B strand and F strand. IgSF domains can be divided into 4 main classes based on their structures and sequences: the Variable (V), Constant 1 (C1), Constant 2 (C2), and Intermediate (I) sets. The second Ig-like domain of JAM1 is a member of the I-set Ig domains, having A-B-E-D strands in one beta-sheet and A'-G-F-C-C' in the other. Like the V-set Ig domains, the A strand of the I-set is discontinuous but lacks a C" strand. I-set domains are found in several cell adhesion molecules (such as VCAM, ICAM, and MADCAM), and are also present in numerous other diverse protein families, including several tyrosine-protein kinase receptors. Pssm-ID: 409542 Cd Length: 97 Bit Score: 38.84 E-value: 1.67e-03
|
|||||||
| IgC_1_Robo | cd07693 | First immunoglobulin (Ig)-like constant domain in Robo (roundabout) receptors, and similar ... |
52-153 | 2.17e-03 | |||
First immunoglobulin (Ig)-like constant domain in Robo (roundabout) receptors, and similar domains; The members here are composed of the first immunoglobulin (Ig)-like domain in Roundabout (Robo) receptors. Robo receptors play a role in the development of the central nervous system (CNS), and are receptors of Slit protein. Slit is a repellant secreted by the neural cells in the midline. Slit acts through Robo to prevent most neurons from crossing the midline from either side. Three mammalian Robo homologs (Robo1, Robo2, and Robo3), and three mammalian Slit homologs (Slit1, Slit2, Slit3), have been identified. Commissural axons, which cross the midline, express low levels of Robo; longitudinal axons, which avoid the midline, express high levels of Robo. Robo1, Robo2, and Robo3 are expressed by commissural neurons in the vertebrate spinal cord and Slit1, Slit2,and Slit3 are expressed at the ventral midline. Robo3 is a divergent member of the Robo family which instead of being a positive regulator of Slit responsiveness, antagonizes Slit responsiveness in precrossing axons. The Slit-Robo interaction is mediated by the second leucine-rich repeat (LRR) domain of Slit and the two N-terminal Ig domains of Robo, Ig1 and Ig2. The primary Robo binding site for Slit2 has been shown by surface plasmon resonance experiments and mutational analysis to be is the Ig1 domain, while the Ig2 domain has been proposed to harbor a weak secondary binding site. Pssm-ID: 409490 [Multi-domain] Cd Length: 99 Bit Score: 38.30 E-value: 2.17e-03
|
|||||||
| IgI_3_WFIKKN-like | cd05765 | Third immunoglobulin-like domain of the human WFIKKN (WAP, follistatin, immunoglobulin, Kunitz ... |
162-233 | 2.37e-03 | |||
Third immunoglobulin-like domain of the human WFIKKN (WAP, follistatin, immunoglobulin, Kunitz and NTR domain-containing protein), and similar domains; member of the I-set of Ig superfamily (IgSF) domains; The members here are composed of the third immunoglobulin-like domain of the human WFIKKN (WAP, follistatin, immunoglobulin, Kunitz and NTR domain-containing protein) and similar proteins. WFIKKN is a secreted protein that consists of multiple types of protease inhibitory modules, including two tandem Kunitz-type protease inhibitor-domains. The Ig superfamily is a heterogenous group of proteins built on a common fold comprised of a sandwich of two beta sheets. Members of the Ig superfamily are components of immunoglobulin, neuroglia, cell surface glycoproteins, such as T-cell receptors, CD2, CD4, CD8, and membrane glycoproteins, such as butyrophilin and chondroitin sulfate proteoglycan core protein. A predominant feature of most Ig domains is a disulfide bridge connecting the two beta-sheets with a tryptophan residue packed against the disulfide bond. This group belongs to the I-set of IgSF domains, having A-B-E-D strands in one beta-sheet and A'-G-F-C-C' in the other. Like the V-set Ig domains, members of the I-set have a discontinuous A strand, but lack a C" strand. Pssm-ID: 409422 [Multi-domain] Cd Length: 95 Bit Score: 38.30 E-value: 2.37e-03
|
|||||||
| IgI_Perlecan_like | cd05754 | Immunoglobulin (Ig)-like domain found in Perlecan and similar proteins; member of the I-set of ... |
161-233 | 2.39e-03 | |||
Immunoglobulin (Ig)-like domain found in Perlecan and similar proteins; member of the I-set of Ig superfamily (IgSF) domains; The members here are composed of the third immunoglobulin (Ig)-like domain found in Perlecan. Perlecan is a large multi-domain heparin sulfate proteoglycan, important in tissue development and organogenesis. Perlecan can be represented as 5 major portions; its fourth major portion (domain IV) is a tandem repeat of immunoglobulin-like domains (Ig2-Ig15) which can vary in size due to alternative splicing. Perlecan binds many cellular and extracellular ligands. Its domain IV region has many binding sites. Some of these have been mapped at the level of individual Ig-like domains, including a site restricted to the Ig5 domain for heparin/sulfatide, a site restricted to the Ig3 domain for nidogen-1 and nidogen-2, a site restricted to Ig4-5 for fibronectin, and sites restricted to Ig2 and to Ig13-15 for fibulin-2. This group belongs to the I-set of IgSF domains, having A-B-E-D strands in one beta-sheet and A'-G-F-C-C' in the other. Like the V-set Ig domains, members of the I-set have a discontinuous A strand, but lack a C" strand. Pssm-ID: 409412 Cd Length: 85 Bit Score: 37.92 E-value: 2.39e-03
|
|||||||
| IgC2_3_Dscam | cd20957 | Third immunoglobulin domain of the Drosophila melanogaster Dscam protein, and similar domains; ... |
162-233 | 2.72e-03 | |||
Third immunoglobulin domain of the Drosophila melanogaster Dscam protein, and similar domains; a member of the Constant 2 (C2)-set of IgSF domains; The members here are composed of the third immunoglobulin domain of the Drosophila melanogaster Down syndrome cell adhesion molecule (DSCAM) protein and similar proteins. Down syndrome cell adhesion molecule (DSCAM) is a cell adhesion molecule that plays critical roles in neural development, including axon guidance and branching, axon target recognition, self-avoidance and synaptic formation. DSCAM belongs to the immunoglobulin superfamily and contributes to defects in the central nervous system in Down syndrome patients. Vertebrate DSCAMs differ from Drosophila Dscam1 in that they lack the extensive alternative splicing that occurs in the insect gene. Drosophila melanogaster Dscam has 38,016 isoforms generated by the alternative splicing of four variable exon clusters, which allows every neuron in the fly to display a distinctive set of Dscam proteins on its cell surface. Drosophila Dscam1 is a cell-surface protein that plays important roles in neural development and axon tiling of neurons. It is shown that thousands of isoforms bind themselves through specific homophilic (self-binding) interactions, a process which mediates cellular self-recognition. Drosophila Dscam2 is also alternatively spliced and plays a key role in the development of two visual system neurons, monopolar cells L1 and L2. IgSF domains can be divided into 4 main classes based on their structures and sequences: the Variable (V), Constant 1 (C1), Constant 2 (C2), and Intermediate (I) sets. This group belongs to the C2-set of IgSF domains, having A, B, and E strands in one beta-sheet and A', G, F, C, and C' in the other. Unlike other Ig domain sets, the C2-set lacks the D strand. Pssm-ID: 409549 [Multi-domain] Cd Length: 88 Bit Score: 37.90 E-value: 2.72e-03
|
|||||||
| Ig1_IL1R_like | cd05756 | First immunoglobulin (Ig)-like domain of interleukin-1 receptor (IL1R), and similar domains; ... |
191-246 | 2.89e-03 | |||
First immunoglobulin (Ig)-like domain of interleukin-1 receptor (IL1R), and similar domains; The members here are composed of the first immunoglobulin (Ig)-like domain of interleukin-1 receptor (IL1R; also known as cluster of differentiation (CD) 121). IL-1 alpha and IL-1 beta are cytokines which participate in the regulation of inflammation, immune responses, and hematopoiesis. These cytokines bind to the IL-1 receptor type 1 (IL1R1), which is activated on additional association with interleukin-1 receptor accessory protein (IL1RAP). IL-1 also binds a second receptor designated type II (IL1R2). Mature IL1R1 consists of three Ig-like domains, a transmembrane domain, and a large cytoplasmic domain. Mature IL1R2 is organized similarly except that it has a short cytoplasmic domain. The latter does not initiate signal transduction. A naturally occurring cytokine IL-1RA (IL-1 receptor antagonist) is widely expressed and binds to IL-1 receptors, inhibiting the binding of IL-1 alpha and IL-1 beta. Pssm-ID: 409414 Cd Length: 96 Bit Score: 38.17 E-value: 2.89e-03
|
|||||||
| Death | cd01670 | Death Domain: a protein-protein interaction domain; Death Domains (DDs) are protein-protein ... |
870-926 | 2.96e-03 | |||
Death Domain: a protein-protein interaction domain; Death Domains (DDs) are protein-protein interaction domains found in a variety of domain architectures. Their common feature is that they form homodimers by self-association or heterodimers by associating with other members of the DD superfamily including CARD (Caspase activation and recruitment domain), DED (Death Effector Domain), and PYRIN. Structural analysis of DD-DD complexes show that the domains interact with each other in many different ways. DD-containing proteins serve as adaptors in signaling pathways and they can recruit other proteins into signaling complexes. In mammals, they are prominent components of the programmed cell death (apoptosis) pathway and are found in a number of other signaling pathways. In invertebrates, they are involved in transcriptional regulation of zygotic patterning genes in insect embryogenesis, and are components of the ToII/NF-kappaB pathway, a conserved innate immune pathway in animal cells. Pssm-ID: 260017 [Multi-domain] Cd Length: 79 Bit Score: 37.26 E-value: 2.96e-03
|
|||||||
| Ig2_FcgammaR_like | cd05753 | Second immunoglobulin (Ig)-like domain of Fcgamma-receptors (FcgammaRs), and similar domains; ... |
174-249 | 5.14e-03 | |||
Second immunoglobulin (Ig)-like domain of Fcgamma-receptors (FcgammaRs), and similar domains; The members here are composed of the second immunoglobulin (Ig)-like domain of Fcgamma-receptors (FcgammaRs). Interactions between IgG and FcgammaR are important to the initiation of cellular and humoral response. IgG binding to FcgammaR leads to a cascade of signals and ultimately to functions such as antibody-dependent-cellular-cytotoxicity (ADCC), endocytosis, phagocytosis, release of inflammatory mediators, etc. FcgammaR has two Ig-like domains. This group also contains FcepsilonRI which binds IgE with high affinity. Pssm-ID: 409411 Cd Length: 83 Bit Score: 36.90 E-value: 5.14e-03
|
|||||||
| Blast search parameters | ||||
|
