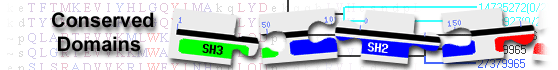Exonuclease-Endonuclease-Phosphatase (EEP) domain superfamily; This large superfamily includes ...
199-530
0e+00
Exonuclease-Endonuclease-Phosphatase (EEP) domain superfamily; This large superfamily includes the catalytic domain (exonuclease/endonuclease/phosphatase or EEP domain) of a diverse set of proteins including the ExoIII family of apurinic/apyrimidinic (AP) endonucleases, inositol polyphosphate 5-phosphatases (INPP5), neutral sphingomyelinases (nSMases), deadenylases (such as the vertebrate circadian-clock regulated nocturnin), bacterial cytolethal distending toxin B (CdtB), deoxyribonuclease 1 (DNase1), the endonuclease domain of the non-LTR retrotransposon LINE-1, and related domains. These diverse enzymes share a common catalytic mechanism of cleaving phosphodiester bonds; their substrates range from nucleic acids to phospholipids and perhaps proteins.
The actual alignment was detected with superfamily member cd09099:
Pssm-ID: 469791 Cd Length: 336 Bit Score: 744.53 E-value: 0e+00
Catalytic inositol polyphosphate 5-phosphatase (INPP5c) domain of synaptojanin 2; This ...
199-530
0e+00
Catalytic inositol polyphosphate 5-phosphatase (INPP5c) domain of synaptojanin 2; This subfamily contains the INPP5c domains of human synaptojanin 2 (Synj2) and related proteins. It belongs to a family of Mg2+-dependent inositol polyphosphate 5-phosphatases, which hydrolyze the 5-phosphate from the inositol ring of various 5-position phosphorylated phosphoinositides (PIs) and inositol phosphates (IPs), and to the large EEP (exonuclease/endonuclease/phosphatase) superfamily that contains functionally diverse enzymes that share a common catalytic mechanism of cleaving phosphodiester bonds. Synj2 can hydrolyze phosphatidylinositol diphosphate (PIP2) to phosphatidylinositol phosphate (PIP). In addition to this INPP5c domain, these proteins contain an N-terminal Sac1-like domain; the Sac1 domain can dephosphorylate a variety of phosphoinositides in vitro. Synj2 occurs as multiple alternative splice variants in various tissues. These variants share the INPP5c domain and the Sac1 domain. Synj2A is recruited to the mitochondria via its interaction with OMP25, a mitochondrial outer membrane protein. Synj2B is found at nerve terminals in the brain and at the spermatid manchette in testis. Synj2B undergoes further alternative splicing to give 2B1 and 2B2. In clathrin-mediated endocytosis, Synj2 participates in the formation of clathrin-coated pits, and perhaps also in vesicle decoating. Rac1 GTPase regulates the intracellular localization of Synj2 forms, but not Synj1. Synj2 may contribute to the role of Rac1 in cell migration and invasion, and is a potential target for therapeutic intervention in malignant tumors.
Pssm-ID: 197333 Cd Length: 336 Bit Score: 744.53 E-value: 0e+00
RNA recognition motif (RRM) found in synaptojanin-2 and similar proteins; This subgroup ...
558-635
2.25e-46
RNA recognition motif (RRM) found in synaptojanin-2 and similar proteins; This subgroup corresponds to the RRM of synaptojanin-2, also termed synaptic inositol-1,4,5-trisphosphate 5-phosphatase 2, an ubiquitously expressed central regulatory enzyme in the phosphoinositide-signaling cascade. As a novel Rac1 effector regulating the early step of clathrin-mediated endocytosis, synaptojanin-2 acts as a polyphosphoinositide phosphatase directly and specifically interacting with Rac1 in a GTP-dependent manner. It mediates the inhibitory effect of Rac1 on endocytosis and plays an important role in the Rac1-mediated control of cell growth. Synaptojanin-2 shows high sequence homology to the N-terminal Sac1p homology domain, the central inositol 5-phosphatase domain, the putative RNA recognition motif (RRM) of synaptojanin-1, but differs in the proline-rich region.
Pssm-ID: 410119 [Multi-domain] Cd Length: 78 Bit Score: 160.33 E-value: 2.25e-46
Catalytic inositol polyphosphate 5-phosphatase (INPP5c) domain of synaptojanin 2; This ...
199-530
0e+00
Catalytic inositol polyphosphate 5-phosphatase (INPP5c) domain of synaptojanin 2; This subfamily contains the INPP5c domains of human synaptojanin 2 (Synj2) and related proteins. It belongs to a family of Mg2+-dependent inositol polyphosphate 5-phosphatases, which hydrolyze the 5-phosphate from the inositol ring of various 5-position phosphorylated phosphoinositides (PIs) and inositol phosphates (IPs), and to the large EEP (exonuclease/endonuclease/phosphatase) superfamily that contains functionally diverse enzymes that share a common catalytic mechanism of cleaving phosphodiester bonds. Synj2 can hydrolyze phosphatidylinositol diphosphate (PIP2) to phosphatidylinositol phosphate (PIP). In addition to this INPP5c domain, these proteins contain an N-terminal Sac1-like domain; the Sac1 domain can dephosphorylate a variety of phosphoinositides in vitro. Synj2 occurs as multiple alternative splice variants in various tissues. These variants share the INPP5c domain and the Sac1 domain. Synj2A is recruited to the mitochondria via its interaction with OMP25, a mitochondrial outer membrane protein. Synj2B is found at nerve terminals in the brain and at the spermatid manchette in testis. Synj2B undergoes further alternative splicing to give 2B1 and 2B2. In clathrin-mediated endocytosis, Synj2 participates in the formation of clathrin-coated pits, and perhaps also in vesicle decoating. Rac1 GTPase regulates the intracellular localization of Synj2 forms, but not Synj1. Synj2 may contribute to the role of Rac1 in cell migration and invasion, and is a potential target for therapeutic intervention in malignant tumors.
Pssm-ID: 197333 Cd Length: 336 Bit Score: 744.53 E-value: 0e+00
Catalytic inositol polyphosphate 5-phosphatase (INPP5c) domain of synaptojanins; This ...
199-530
0e+00
Catalytic inositol polyphosphate 5-phosphatase (INPP5c) domain of synaptojanins; This subfamily contains the INPP5c domains of two human synaptojanins, synaptojanin 1 (Synj1) and synaptojanin 2 (Synj2), and related proteins. It belongs to a family of Mg2+-dependent inositol polyphosphate 5-phosphatases, which hydrolyze the 5-phosphate from the inositol ring of various 5-position phosphorylated phosphoinositides (PIs) and inositol phosphates (IPs). They belong to the large EEP (exonuclease/endonuclease/phosphatase) superfamily that contains functionally diverse enzymes that share a common catalytic mechanism of cleaving phosphodiester bonds. Synj1 occurs as two main isoforms: a brain enriched 145 KDa protein (Synj1-145) and a ubiquitously expressed 170KDa protein (Synj1-170). Synj1-145 participates in clathrin-mediated endocytosis. The primary substrate of the Synj1-145 INPP5c domain is PI(4,5)P2, which it converts to PI4P. Synj1-145 may work with membrane curvature sensors/generators (such as endophilin) to remove PI(4,5)P2 from curved membranes. The recruitment of the INPP5c domain of Synj1-145 to endophilin-induced membranes leads to a fragmentation and condensation of these structures. The PI(4,5)P2 to PI4P conversion may cooperate with dynamin to produce membrane fission. In addition to this INPP5c domain, Synjs contain an N-terminal Sac1-like domain; the Sac1 domain can dephosphorylate a variety of phosphoinositides in vitro. Synj2 can hydrolyze phosphatidylinositol diphosphate (PIP2) to phosphatidylinositol phosphate (PIP). Synj2 occurs as multiple alternative splice variants in various tissues. These variants share the INPP5c domain and the Sac1 domain. Synj2A is recruited to the mitochondria via its interaction with OMP25 (a mitochondrial outer membrane protein). Synj2B is found at nerve terminals in the brain and at the spermatid manchette in testis. Synj2B undergoes further alternative splicing to give 2B1 and 2B2. In clathrin-mediated endocytosis, Synj2 participates in the formation of clathrin-coated pits, and perhaps also in vesicle decoating. Rac1 GTPase regulates the intracellular localization of Synj2 forms, but not Synj1. Synj2 may contribute to the role of Rac1 in cell migration and invasion, and is a potential target for therapeutic intervention in malignant tumors.
Pssm-ID: 197323 [Multi-domain] Cd Length: 328 Bit Score: 637.13 E-value: 0e+00
Catalytic inositol polyphosphate 5-phosphatase (INPP5c) domain of synaptojanin 1; This ...
199-530
0e+00
Catalytic inositol polyphosphate 5-phosphatase (INPP5c) domain of synaptojanin 1; This subfamily contains the INPP5c domains of human synaptojanin 1 (Synj1) and related proteins. It belongs to a family of Mg2+-dependent inositol polyphosphate 5-phosphatases, which hydrolyze the 5-phosphate from the inositol ring of various 5-position phosphorylated phosphoinositides (PIs) and inositol phosphates (IPs), and to the large EEP (exonuclease/endonuclease/phosphatase) superfamily that contains functionally diverse enzymes that share a common catalytic mechanism of cleaving phosphodiester bonds. Synj1 occurs as two main isoforms: a brain enriched 145 KDa protein (Synj1-145) and a ubiquitously expressed 170KDa protein (Synj1-170). Synj1-145 participates in clathrin-mediated endocytosis. The primary substrate of the Synj1-145 INPP5c domain is PI(4,5)P2, which it converts to PI4P. Synj1-145 may work with membrane curvature sensors/generators (such as endophilin) to remove PI(4,5)P2 from curved membranes. The recruitment of the INPP5c domain of Synj1-145 to endophilin-induced membranes leads to a fragmentation and condensation of these structures. The PI(4,5)P2 to PI4P conversion may cooperate with dynamin to produce membrane fission. In addition to this INPP5c domain, these proteins contain an N-terminal Sac1-like domain; the Sac1 domain can dephosphorylate a variety of phosphoinositides in vitro.
Pssm-ID: 197332 Cd Length: 336 Bit Score: 535.39 E-value: 0e+00
Catalytic domain of inositol polyphosphate 5-phosphatases; Inositol polyphosphate ...
199-530
1.11e-123
Catalytic domain of inositol polyphosphate 5-phosphatases; Inositol polyphosphate 5-phosphatases (5-phosphatases) are signal-modifying enzymes, which hydrolyze the 5-phosphate from the inositol ring of specific 5-position phosphorylated phosphoinositides (PIs) and inositol phosphates (IPs), such as PI(4,5)P2, PI(3,4,5)P3, PI(3,5)P2, I(1,4,5)P3, and I(1,3,4,5)P4. These enzymes are Mg2+-dependent, and belong to the large EEP (exonuclease/endonuclease/phosphatase) superfamily that contains functionally diverse enzymes that share a common catalytic mechanism of cleaving phosphodiester bonds. In addition to this INPP5c domain, 5-phosphatases often contain additional domains and motifs, such as the SH2 domain, the Sac-1 domain, the proline-rich domain (PRD), CAAX, RhoGAP (RhoGTPase-activating protein), and SKICH [SKIP (skeletal muscle- and kidney-enriched inositol phosphatase) carboxyl homology] domains, that are important for protein-protein interactions and/or for the subcellular localization of these enzymes. 5-phosphatases incorporate into large signaling complexes, and regulate diverse cellular processes including postsynaptic vesicular trafficking, insulin signaling, cell growth and survival, and endocytosis. Loss or gain of function of 5-phosphatases is implicated in certain human diseases. This family also contains a functionally unrelated nitric oxide transport protein, Cimex lectularius (bedbug) nitrophorin, which catalyzes a heme-assisted S-nitrosation of a proximal thiolate; the heme however binds at a site distinct from the active site of the 5-phosphatases.
Pssm-ID: 197308 [Multi-domain] Cd Length: 299 Bit Score: 382.07 E-value: 1.11e-123
Catalytic inositol polyphosphate 5-phosphatase (INPP5c) domain of Saccharomyces cerevisiae ...
199-530
1.72e-102
Catalytic inositol polyphosphate 5-phosphatase (INPP5c) domain of Saccharomyces cerevisiae Inp51p, Inp52p, and Inp53p, and related proteins; This subfamily contains the INPP5c domain of three Saccharomyces cerevisiae synaptojanin-like inositol polyphosphate 5-phosphatases (INP51, INP52, and INP53), Schizosaccharomyces pombe synaptojanin (SPsynaptojanin), and related proteins. It belongs to a family of Mg2+-dependent inositol polyphosphate 5-phosphatases, which hydrolyze the 5-phosphate from the inositol ring of various 5-position phosphorylated phosphoinositides (PIs) and inositol phosphates (IPs), and to the large EEP (exonuclease/endonuclease/phosphatase) superfamily that contains functionally diverse enzymes that share a common catalytic mechanism of cleaving phosphodiester bonds. In addition to this INPP5c domain, these proteins have an N-terminal catalytic Sac1-like domain (found in other proteins including the phophoinositide phosphatase Sac1p), and a C-terminal proline-rich domain (PRD). The Sac1 domain allows Inp52p and Inp53p to recognize and dephosphorylate a wider range of substrates including PI3P, PI4P, and PI(3,5)P2. The Sac1 domain of Inp51p is non-functional. Disruption of any two of INP51, INP52, and INP53, in S. cerevisiae leads to abnormal vacuolar and plasma membrane morphology. During hyperosmotic stress, Inp52p and Inp53p localize at actin patches, where they may facilitate the hydrolysis of PI(4,5)P2, and consequently promote actin rearrangement to regulate cell growth. SPsynaptojanin is also active against a range of soluble and lipid inositol phosphates, including I(1,4,5)P3, I(1,3,4,5)P4, I(1,4,5,6)P4, PI(4,5)P2, and PIP3. Transformation of S. cerevisiae with a plasmid expressing the SPsynaptojanin 5-phosphatase domain rescues inp51/inp52/inp53 triple-mutant strains.
Pssm-ID: 197324 Cd Length: 291 Bit Score: 325.45 E-value: 1.72e-102
Catalytic inositol polyphosphate 5-phosphatase (INPP5c) domain of Type II inositol ...
199-529
3.45e-94
Catalytic inositol polyphosphate 5-phosphatase (INPP5c) domain of Type II inositol polyphosphate 5-phosphatase I, Oculocerebrorenal syndrome of Lowe 1, and related proteins; This subfamily contains the INPP5c domain of type II inositol polyphosphate 5-phosphatase I (INPP5B), Oculocerebrorenal syndrome of Lowe 1 (OCRL-1), and related proteins. It belongs to a family of Mg2+-dependent inositol polyphosphate 5-phosphatases, which hydrolyze the 5-phosphate from the inositol ring of various 5-position phosphorylated phosphoinositides (PIs) and inositol phosphates (IPs), and to the large EEP (exonuclease/endonuclease/phosphatase) superfamily that contains functionally diverse enzymes that share a common catalytic mechanism of cleaving phosphodiester bonds. INPP5B and OCRL1 preferentially hydrolyze the 5-phosphate of phosphatidylinositol (4,5)- bisphosphate [PI(4,5)P2] and phosphatidylinositol (3,4,5)- trisphosphate [PI(3,4,5)P3]. INPP5B can also hydrolyze soluble inositol (1,4,5)-trisphosphate [I(1,4,5)P3] and inositol (1,3,4,5)-tetrakisphosphate [I(1,3,4,5)P4]. INPP5B participates in the endocytic pathway and in the early secretory pathway. In the latter, it may function in retrograde ERGIC (ER-to-Golgi intermediate compartment)-to-ER transport; it binds specific RAB proteins within the secretory pathway. In the endocytic pathway, it binds RAB5 and during endocytosis, may function in a RAB5-controlled cascade for converting PI(3,4,5)P3 to phosphatidylinositol 3-phosphate (PI3P). This cascade may link growth factor signaling and membrane dynamics. Mutation in OCRL1 is implicated in Lowe syndrome, an X-linked recessive multisystem disorder, which includes defects in eye, brain, and kidney function, and in Type 2 Dent's disease, a disorder with only the renal symptoms. OCRL-1 may have a role in membrane trafficking within the endocytic pathway and at the trans-Golgi network, and may participate in actin dynamics or signaling from endomembranes. OCRL1 and INPP5B have overlapping functions: deletion of both 5-phosphatases in mice is embryonic lethal, deletion of OCRL1 alone has no phenotype, and deletion of Inpp5b alone has only a mild phenotype (male sterility). Several of the proteins that interact with OCRL1 also bind INPP5B, for examples, inositol polyphosphate phosphatase interacting protein of 27kDa (IPIP27)A and B (also known as Ses1 and 2), and endocytic signaling adaptor APPL1. OCRL1, but not INPP5B, binds clathrin heavy chain, the plasma membrane AP2 adaptor subunit alpha-adaptin. In addition to this INPP5c domain, most proteins in this subfamily have a C-terminal RhoGAP (GTPase-activator protein [GAP] for Rho-like small GTPases) domain.
Pssm-ID: 197327 Cd Length: 292 Bit Score: 303.08 E-value: 3.45e-94
Catalytic inositol polyphosphate 5-phosphatase (INPP5c) domain of inositol polyphosphate ...
241-530
9.32e-70
Catalytic inositol polyphosphate 5-phosphatase (INPP5c) domain of inositol polyphosphate 5-phosphatase J and related proteins; INPP5c domain of Inositol polyphosphate-5-phosphatase J (INPP5J), also known as PIB5PA or PIPP, and related proteins. This subfamily belongs to a family of Mg2+-dependent inositol polyphosphate 5-phosphatases, which hydrolyze the 5-phosphate from the inositol ring of various 5-position phosphorylated phosphoinositides (PIs) and inositol phosphates (IPs), and to the large EEP (exonuclease/endonuclease/phosphatase) superfamily that contains functionally diverse enzymes that share a common catalytic mechanism of cleaving phosphodiester bonds. INPP5J hydrolyzes PI(4,5)P2, I(1,4,5)P3, and I(1,3,4,5)P4 at ruffling membranes. These proteins contain a C-terminal, SKIP carboxyl homology domain (SKICH), which may direct plasma membrane ruffle localization.
Pssm-ID: 197328 Cd Length: 300 Bit Score: 235.73 E-value: 9.32e-70
Catalytic inositol polyphosphate 5-phosphatase (INPP5c) domain of Inositol ...
197-530
1.24e-60
Catalytic inositol polyphosphate 5-phosphatase (INPP5c) domain of Inositol polyphosphate-5-phosphatase E and related proteins; INPP5c domain of Inositol polyphosphate-5-phosphatase E (also called type IV or 72 kDa 5-phosphatase), rat pharbin, and related proteins. This subfamily belongs to a family of Mg2+-dependent inositol polyphosphate 5-phosphatases, which hydrolyze the 5-phosphate from the inositol ring of various 5-position phosphorylated phosphoinositides (PIs) and inositol phosphates (IPs), and to the large EEP (exonuclease/endonuclease/phosphatase) superfamily that contains functionally diverse enzymes that share a common catalytic mechanism of cleaving phosphodiester bonds. INPP5E hydrolyzes the 5-phosphate from PI(3,5)P2, PI(4,5)P2 and PI(3,4,5)P3, forming PI3P, PI4P, and PI(3,4)P2, respectively. It is a very potent PI(3,4,5)P3 5-phosphatase. Its intracellular localization is chiefly cytosolic, with pronounced perinuclear/Golgi localization. INPP5E also has an N-terminal proline rich domain (PRD) and a C-terminal CAAX motif. This protein is expressed in a variety of tissues, including the breast, brain, testis, and haemopoietic cells. It is differentially expressed in several cancers, for example, it is up-regulated in cervical cancer and down-regulated in stomach cancer. It is a candidate target for therapeutics of obesity and related disorders, as it is expressed in the hypothalamus, and following insulin stimulation, it undergoes tyrosine phosphorylation, associates with insulin receptor substrate-1, -2, and PI3-kinase, and become active as a 5-phosphatase. INPP5E may play a role, along with other 5-phosphatases SHIP2 and SKIP, in regulating glucose homoeostasis and energy metabolism. Mice deficient in INPPE5 develop a multi-organ disorder associated with structural defects of the primary cilium.
Pssm-ID: 197329 Cd Length: 298 Bit Score: 209.97 E-value: 1.24e-60
RNA recognition motif (RRM) found in synaptojanin-2 and similar proteins; This subgroup ...
558-635
2.25e-46
RNA recognition motif (RRM) found in synaptojanin-2 and similar proteins; This subgroup corresponds to the RRM of synaptojanin-2, also termed synaptic inositol-1,4,5-trisphosphate 5-phosphatase 2, an ubiquitously expressed central regulatory enzyme in the phosphoinositide-signaling cascade. As a novel Rac1 effector regulating the early step of clathrin-mediated endocytosis, synaptojanin-2 acts as a polyphosphoinositide phosphatase directly and specifically interacting with Rac1 in a GTP-dependent manner. It mediates the inhibitory effect of Rac1 on endocytosis and plays an important role in the Rac1-mediated control of cell growth. Synaptojanin-2 shows high sequence homology to the N-terminal Sac1p homology domain, the central inositol 5-phosphatase domain, the putative RNA recognition motif (RRM) of synaptojanin-1, but differs in the proline-rich region.
Pssm-ID: 410119 [Multi-domain] Cd Length: 78 Bit Score: 160.33 E-value: 2.25e-46
Catalytic inositol polyphosphate 5-phosphatase (INPP5c) domain of SH2 domain containing inositol polyphosphate 5-phosphatase-1 and -2, and related proteins; This subfamily contains the INPP5c domain of SHIP1 (SH2 domain containing inositol polyphosphate 5-phosphatase-1, also known as SHIP/INPP5D), and SHIP2 (also known as INPPL1). It belongs to a family of Mg2+-dependent inositol polyphosphate 5-phosphatases, which hydrolyze the 5-phosphate from the inositol ring of various 5-position phosphorylated phosphoinositides (PIs) and inositol phosphates (IPs), and to the large EEP (exonuclease/endonuclease/phosphatase) superfamily that contains functionally diverse enzymes that share a common catalytic mechanism of cleaving phosphodiester bonds. Both SHIP1 and -2 catalyze the dephosphorylation of the PI, phosphatidylinositol 3,4,5-trisphosphate [PI(3,4,5)P3], to phosphatidylinositol 3,4-bisphosphate [PI(3,4)P2]. SHIP1 also converts inositol-1,3,4,5- polyphosphate [I(1,3,4,5)P4] to inositol-1,3,4-polyphosphate [I(1,3,4)P3]. SHIP1 and SHIP2 have little overlap in their in vivo functions. SHIP1 is a negative regulator of cell growth and plays a major part in mediating the inhibitory signaling in B cells; it is predominantly expressed in hematopoietic cells. SHIP2 is as an inhibitor of the insulin signaling pathway, and is implicated in actin structure remodeling, cell adhesion and cell spreading, receptor endocytosis and degradation, and in the JIP1-mediated JNK pathway. SHIP2 is widely expressed, most prominently in brain, heart and in skeletal muscle. In addition to this INPP5c domain, SHIP1 has an N-terminal SH2 domain, two NPXY motifs, and a C-terminal proline-rich region (PRD), while SHIP2 has an N-terminal SH2 domain, a C-terminal proline-rich domain (PRD), which includes a WW-domain binding motif (PPLP), an NPXY motif, and a sterile alpha motif (SAM) domain. The gene encoding SHIP2 is a candidate gene for conferring a predisposition for type 2 diabetes.
Pssm-ID: 197325 Cd Length: 307 Bit Score: 160.11 E-value: 3.04e-43
Catalytic inositol polyphosphate 5-phosphatase (INPP5c) domain of SH2 domain containing inositol polyphosphate 5-phosphatase-1 and related proteins; This subfamily contains the INPP5c domain of SHIP1 (SH2 domain containing inositol polyphosphate 5-phosphatase-1, also known as SHIP/INPP5D) and related proteins. It belongs to a family of Mg2+-dependent inositol polyphosphate 5-phosphatases, which hydrolyze the 5-phosphate from the inositol ring of various 5-position phosphorylated phosphoinositides (PIs) and inositol phosphates (IPs), and to the large EEP (exonuclease/endonuclease/phosphatase) superfamily that contains functionally diverse enzymes that share a common catalytic mechanism of cleaving phosphodiester bonds. SHIP1's enzymic activity is restricted to phosphatidylinositol 3,4,5-trisphosphate [PI (3,4,5)P3] and inositol-1,3,4,5- polyphosphate [I(1,3,4,5)P4]. It converts these two phosphoinositides to phosphatidylinositol 3,4-bisphosphate [PI (3,4)P2] and inositol-1,3,4-polyphosphate [I(1,3,4)P3], respectively. SHIP1 is a negative regulator of cell growth and plays a major part in mediating the inhibitory signaling in B cells; it is predominantly expressed in hematopoietic cells. In addition to this INPP5c domain, SHIP1 has an N-terminal SH2 domain, two NPXY motifs, and a C-terminal proline-rich region (PRD). SHIP1's phosphorylated NPXY motifs interact with proteins with phosphotyrosine binding (PTB) domains, and facilitate the translocation of SHIP1 to the plasma membrane to hydrolyze PI(3,4,5)P3. SHIP1 generally acts to oppose the activity of phosphatidylinositol 3-kinase (PI3K). It acts as a negative signaling molecule, reducing the levels of PI(3,4,5)P3, thereby removing the latter as a membrane-targeting signal for PH domain-containing effector molecules. SHIP1 may also, in certain contexts, amplify PI3K signals. SHIP1 and SHIP2 have little overlap in their in vivo functions.
Pssm-ID: 197334 Cd Length: 307 Bit Score: 159.00 E-value: 7.15e-43
Catalytic inositol polyphosphate 5-phosphatase (INPP5c) domain of SH2 domain containing inositol 5-phosphatase-2 and related proteins; This subfamily contains the INPP5c domain of SHIP2 (SH2 domain containing inositol 5-phosphatase-2, also called INPPL1) and related proteins. It belongs to a family of Mg2+-dependent inositol polyphosphate 5-phosphatases, which hydrolyze the 5-phosphate from the inositol ring of various 5-position phosphorylated phosphoinositides (PIs) and inositol phosphates (IPs), and to the large EEP (exonuclease/endonuclease/phosphatase) superfamily that contains functionally diverse enzymes that share a common catalytic mechanism of cleaving phosphodiester bonds. SHIP2 catalyzes the dephosphorylation of the PI, phosphatidylinositol 3,4,5-trisphosphate [PI(3,4,5)P3], to phosphatidylinositol 3,4-bisphosphate [PI(3,4)P2]. SHIP2 is widely expressed, most prominently in brain, heart and in skeletal muscle. SHIP2 is an inhibitor of the insulin signaling pathway. It is implicated in actin structure remodeling, cell adhesion and cell spreading, receptor endocytosis and degradation, and in the JIP1-mediated JNK pathway. Its interacting partners include filamin/actin, p130Cas, Shc, Vinexin, Interesectin 1, and c-Jun NH2-terminal kinase (JNK)-interacting protein 1 (JIP1). A large variety of extracellular stimuli appear to lead to the tyrosine phosphorylation of SHIP2, including epidermal growth factor (EGF), platelet-derived growth factor (PDGF), insulin, macrophage colony-stimulating factor (M-CSF) and hepatocyte growth factor (HGF). SHIP2 is localized to the cytosol in quiescent cells; following growth factor stimulation and /or cell adhesion, it relocalizes to membrane ruffles. In addition to this INPP5c domain, SHIP2 has an N-terminal SH2 domain, a C-terminal proline-rich domain (PRD), which includes a WW-domain binding motif (PPLP), an NPXY motif and a sterile alpha motif (SAM) domain. The gene encoding SHIP2 is a candidate for conferring a predisposition for type 2 diabetes; it has been suggested that suppression of SHIP2 may be of benefit in the treatment of obesity and thereby prevent type 2 diabetes. SHIP2 and SHIP1 have little overlap in their in vivo functions.
Pssm-ID: 197335 Cd Length: 304 Bit Score: 157.06 E-value: 3.30e-42
RNA recognition motif (RRM) found in synaptojanin-1, synaptojanin-2 and similar proteins; This ...
558-635
3.18e-32
RNA recognition motif (RRM) found in synaptojanin-1, synaptojanin-2 and similar proteins; This subfamily corresponds to the RRM of two active phosphatidylinositol phosphate phosphatases, synaptojanin-1 and synaptojanin-2. They have different interaction partners and are likely to have different biological functions. Synaptojanin-1 was originally identified as one of the major Grb2-binding proteins that may participate in synaptic vesicle endocytosis. It also acts as a Src homology 3 (SH3) domain-binding brain-specific inositol 5-phosphatase with a putative role in clathrin-mediated endocytosis. Synaptojanin-2 is a ubiquitously expressed homolog of synaptojanin-1. It is a novel Rac1 effector regulating the early step of clathrin-mediated endocytosis. Synaptojanin-2 directly and specifically interacts with Rac1 in a GTP-dependent manner. It mediates the inhibitory effect of Rac1 on endocytosis and plays an important role in the Rac1-mediated control of cell growth. Both, synaptojanin-1 and synaptojanin-2, have two tissue-specific alternative splicing isoforms, a shorter isoform expressed in brain and a longer isoform in peripheral tissues. Synaptojanin-1 contains an N-terminal domain homologous to the cytoplasmic portion of the yeast protein Sac1p, a central inositol 5-phosphatase domain followed by a putative RNA recognition motif (RRM), also termed RBD (RNA binding domain) or RNP (ribonucleoprotein domain), and a C-terminal proline-rich region mediating the binding of synaptojanin-1 to various SH3 domain-containing proteins including amphiphysin, SH3p4, SH3p8, SH3p13, and Grb2. Synaptojanin-2 shows high sequence homology to the N-terminal Sac1p homology domain, the central inositol 5-phosphatase domain, the putative RNA recognition motif (RRM) of synaptojanin-1, but differs in the proline-rich region.
Pssm-ID: 409874 [Multi-domain] Cd Length: 77 Bit Score: 119.84 E-value: 3.18e-32
RNA recognition motif (RRM) found in synaptojanin-1 and similar proteins; This subgroup ...
558-635
1.28e-11
RNA recognition motif (RRM) found in synaptojanin-1 and similar proteins; This subgroup corresponds to the RRM of synaptojanin-1, also termed synaptojanin, or synaptic inositol-1,4,5-trisphosphate 5-phosphatase 1, originally identified as one of the major Grb2-binding proteins that may participate in synaptic vesicle endocytosis. It also acts as a Src homology 3 (SH3) domain-binding brain-specific inositol 5-phosphatase with a putative role in clathrin-mediated endocytosis. Synaptojanin-1 contains an N-terminal domain homologous to the cytoplasmic portion of the yeast protein Sac1p, a central inositol 5-phosphatase domain followed by a putative RNA recognition motif (RRM), also termed RBD (RNA binding domain) or RNP (ribonucleoprotein domain), and a C-terminal proline-rich region mediating the binding of synaptojanin-1 to various SH3 domain-containing proteins including amphiphysin, SH3p4, SH3p8, SH3p13, and Grb2. Synaptojanin-1 has two tissue-specific alternative splicing isoforms, synaptojanin-145 expressed in brain and synaptojanin-170 expressed in peripheral tissues. Synaptojanin-145 is very abundant in nerve terminals and may play an essential role in the clathrin-mediated endocytosis of synaptic vesicles. In contrast to synaptojanin-145, synaptojanin-170 contains three unique asparagine-proline-phenylalanine (NPF) motifs in the C-terminal region and may functions as a potential binding partner for Eps15, a clathrin coat-associated protein acting as a major substrate for the tyrosine kinase activity of the epidermal growth factor receptor.
Pssm-ID: 410118 Cd Length: 77 Bit Score: 61.27 E-value: 1.28e-11
Exonuclease-Endonuclease-Phosphatase (EEP) domain superfamily; This large superfamily includes ...
280-529
3.39e-11
Exonuclease-Endonuclease-Phosphatase (EEP) domain superfamily; This large superfamily includes the catalytic domain (exonuclease/endonuclease/phosphatase or EEP domain) of a diverse set of proteins including the ExoIII family of apurinic/apyrimidinic (AP) endonucleases, inositol polyphosphate 5-phosphatases (INPP5), neutral sphingomyelinases (nSMases), deadenylases (such as the vertebrate circadian-clock regulated nocturnin), bacterial cytolethal distending toxin B (CdtB), deoxyribonuclease 1 (DNase1), the endonuclease domain of the non-LTR retrotransposon LINE-1, and related domains. These diverse enzymes share a common catalytic mechanism of cleaving phosphodiester bonds; their substrates range from nucleic acids to phospholipids and perhaps proteins.
Pssm-ID: 197306 [Multi-domain] Cd Length: 241 Bit Score: 64.81 E-value: 3.39e-11
Database: CDSEARCH/cdd Low complexity filter: no Composition Based Adjustment: yes E-value threshold: 0.01
References:
Wang J et al. (2023), "The conserved domain database in 2023", Nucleic Acids Res.51(D)384-8.
Lu S et al. (2020), "The conserved domain database in 2020", Nucleic Acids Res.48(D)265-8.
Marchler-Bauer A et al. (2017), "CDD/SPARCLE: functional classification of proteins via subfamily domain architectures.", Nucleic Acids Res.45(D)200-3.
of the residues that compose this conserved feature have been mapped to the query sequence.
Click on the triangle to view details about the feature, including a multiple sequence alignment
of your query sequence and the protein sequences used to curate the domain model,
where hash marks (#) above the aligned sequences show the location of the conserved feature residues.
The thumbnail image, if present, provides an approximate view of the feature's location in 3 dimensions.
Click on the triangle for interactive 3D structure viewing options.
Functional characterization of the conserved domain architecture found on the query.
Click here to see more details.
This image shows a graphical summary of conserved domains identified on the query sequence.
The Show Concise/Full Display button at the top of the page can be used to select the desired level of detail: only top scoring hits
(labeled illustration) or all hits
(labeled illustration).
Domains are color coded according to superfamilies
to which they have been assigned. Hits with scores that pass a domain-specific threshold
(specific hits) are drawn in bright colors.
Others (non-specific hits) and
superfamily placeholders are drawn in pastel colors.
if a domain or superfamily has been annotated with functional sites (conserved features),
they are mapped to the query sequence and indicated through sets of triangles
with the same color and shade of the domain or superfamily that provides the annotation. Mouse over the colored bars or triangles to see descriptions of the domains and features.
click on the bars or triangles to view your query sequence embedded in a multiple sequence alignment of the proteins used to develop the corresponding domain model.
The table lists conserved domains identified on the query sequence. Click on the plus sign (+) on the left to display full descriptions, alignments, and scores.
Click on the domain model's accession number to view the multiple sequence alignment of the proteins used to develop the corresponding domain model.
To view your query sequence embedded in that multiple sequence alignment, click on the colored bars in the Graphical Summary portion of the search results page,
or click on the triangles, if present, that represent functional sites (conserved features)
mapped to the query sequence.
Concise Display shows only the best scoring domain model, in each hit category listed below except non-specific hits, for each region on the query sequence.
(labeled illustration) Standard Display shows only the best scoring domain model from each source, in each hit category listed below for each region on the query sequence.
(labeled illustration) Full Display shows all domain models, in each hit category below, that meet or exceed the RPS-BLAST threshold for statistical significance.
(labeled illustration) Four types of hits can be shown, as available,
for each region on the query sequence:
specific hits meet or exceed a domain-specific e-value threshold
(illustrated example)
and represent a very high confidence that the query sequence belongs to the same protein family as the sequences use to create the domain model
non-specific hits
meet or exceed the RPS-BLAST threshold for statistical significance (default E-value cutoff of 0.01, or an E-value selected by user via the
advanced search options)
the domain superfamily to which the specific and non-specific hits belong
multi-domain models that were computationally detected and are likely to contain multiple single domains
Retrieve proteins that contain one or more of the domains present in the query sequence, using the Conserved Domain Architecture Retrieval Tool
(CDART).
Modify your query to search against a different database and/or use advanced search options