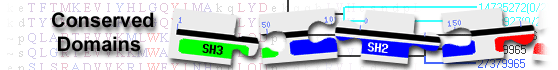


enolase-phosphatase E1 isoform X2 [Mus musculus]
Protein Classification

acireductone synthase( domain architecture ID 11546975)
acireductone synthase is a bifunctional enzyme that catalyzes the enolization of 2,3-diketo-5-methylthiopentyl-1-phosphate (DK-MTP-1-P) into the intermediate 2-hydroxy-3-keto-5-methylthiopentenyl-1-phosphate (HK-MTPenyl-1-P), which is then dephosphorylated to form the acireductone 1,2-dihydroxy-3-keto-5-methylthiopentene (DHK-MTPene)
List of domain hits
| Name | Accession | Description | Interval | E-value | |||
| HAD_EP | cd01629 | Enolase-phosphatase similar to human enolase-phosphatase E1 and and Xanthomonas oryzae pv. ... |
1-146 | 2.70e-87 | |||
Enolase-phosphatase similar to human enolase-phosphatase E1 and and Xanthomonas oryzae pv. Oryzae enolase-phosphatase Xep; Enolase-phosphatase E1 (also called MASA) is a bifunctional enolase- phosphatase which promotes the conversion of 2,3-diketo-5-methylthio-1-phosphopentane to 1,2-dihydroxy-3-keto-5-methylthiopentene anion (an aci-reductone) in the methionine salvage pathway. The catalytic reaction is carried out continuously by enolization and dephosphorylation, and the enolase activity cannot be classified as typical enzymatic enolization. This family belongs to the haloacid dehalogenase-like (HAD) hydrolases, a large superfamily of diverse enzymes that catalyze carbon or phosphoryl group transfer reactions on a range of substrates, using an active site aspartate in nucleophilic catalysis. Members of this superfamily include 2-L-haloalkanoic acid dehalogenase, azetidine hydrolase, phosphonoacetaldehyde hydrolase, phosphoserine phosphatase, phosphomannomutase, P-type ATPases and many others. HAD hydrolases are found in all three kingdoms of life, and most genomes are predicted to contain multiple HAD-like proteins. Members possess a highly conserved alpha/beta core domain, and many also possess a small cap domain, the fold and function of which is variable. HAD hydrolases are sometimes referred to as belonging to the DDDD superfamily of phosphohydrolases. : Pssm-ID: 319768 [Multi-domain] Cd Length: 204 Bit Score: 254.39 E-value: 2.70e-87
|
|||||||
| Name | Accession | Description | Interval | E-value | ||||
| HAD_EP | cd01629 | Enolase-phosphatase similar to human enolase-phosphatase E1 and and Xanthomonas oryzae pv. ... |
1-146 | 2.70e-87 | ||||
Enolase-phosphatase similar to human enolase-phosphatase E1 and and Xanthomonas oryzae pv. Oryzae enolase-phosphatase Xep; Enolase-phosphatase E1 (also called MASA) is a bifunctional enolase- phosphatase which promotes the conversion of 2,3-diketo-5-methylthio-1-phosphopentane to 1,2-dihydroxy-3-keto-5-methylthiopentene anion (an aci-reductone) in the methionine salvage pathway. The catalytic reaction is carried out continuously by enolization and dephosphorylation, and the enolase activity cannot be classified as typical enzymatic enolization. This family belongs to the haloacid dehalogenase-like (HAD) hydrolases, a large superfamily of diverse enzymes that catalyze carbon or phosphoryl group transfer reactions on a range of substrates, using an active site aspartate in nucleophilic catalysis. Members of this superfamily include 2-L-haloalkanoic acid dehalogenase, azetidine hydrolase, phosphonoacetaldehyde hydrolase, phosphoserine phosphatase, phosphomannomutase, P-type ATPases and many others. HAD hydrolases are found in all three kingdoms of life, and most genomes are predicted to contain multiple HAD-like proteins. Members possess a highly conserved alpha/beta core domain, and many also possess a small cap domain, the fold and function of which is variable. HAD hydrolases are sometimes referred to as belonging to the DDDD superfamily of phosphohydrolases. Pssm-ID: 319768 [Multi-domain] Cd Length: 204 Bit Score: 254.39 E-value: 2.70e-87
|
||||||||
| Utr4 | COG4229 | Enolase-phosphatase E1 involved in merthionine salvage [Amino acid transport and metabolism]; |
2-167 | 1.21e-68 | ||||
Enolase-phosphatase E1 involved in merthionine salvage [Amino acid transport and metabolism]; Pssm-ID: 443373 [Multi-domain] Cd Length: 227 Bit Score: 207.70 E-value: 1.21e-68
|
||||||||
| enolase-ppase | TIGR01691 | 2,3-diketo-5-methylthio-1-phosphopentane phosphatase; This enzyme is the enolase-phosphatase ... |
13-167 | 7.53e-61 | ||||
2,3-diketo-5-methylthio-1-phosphopentane phosphatase; This enzyme is the enolase-phosphatase of methionine salvage, a pathway that regenerates methionine from methylthioadenosine (MTA). Adenosylmethionine (AdoMet) is a donor of different moieties for various processes, including methylation reactions. Use of AdoMet for spermidine biosynthesis, which leads to polyamine biosynthesis, leaves MTA as a by-product that must be cleared. In Bacillus subtilis and related species, this single protein is replaced by separate enzymes with enolase and phosphatase activities. [Central intermediary metabolism, Sulfur metabolism] Pssm-ID: 273760 [Multi-domain] Cd Length: 220 Bit Score: 187.74 E-value: 7.53e-61
|
||||||||
| Name | Accession | Description | Interval | E-value | ||||
| HAD_EP | cd01629 | Enolase-phosphatase similar to human enolase-phosphatase E1 and and Xanthomonas oryzae pv. ... |
1-146 | 2.70e-87 | ||||
Enolase-phosphatase similar to human enolase-phosphatase E1 and and Xanthomonas oryzae pv. Oryzae enolase-phosphatase Xep; Enolase-phosphatase E1 (also called MASA) is a bifunctional enolase- phosphatase which promotes the conversion of 2,3-diketo-5-methylthio-1-phosphopentane to 1,2-dihydroxy-3-keto-5-methylthiopentene anion (an aci-reductone) in the methionine salvage pathway. The catalytic reaction is carried out continuously by enolization and dephosphorylation, and the enolase activity cannot be classified as typical enzymatic enolization. This family belongs to the haloacid dehalogenase-like (HAD) hydrolases, a large superfamily of diverse enzymes that catalyze carbon or phosphoryl group transfer reactions on a range of substrates, using an active site aspartate in nucleophilic catalysis. Members of this superfamily include 2-L-haloalkanoic acid dehalogenase, azetidine hydrolase, phosphonoacetaldehyde hydrolase, phosphoserine phosphatase, phosphomannomutase, P-type ATPases and many others. HAD hydrolases are found in all three kingdoms of life, and most genomes are predicted to contain multiple HAD-like proteins. Members possess a highly conserved alpha/beta core domain, and many also possess a small cap domain, the fold and function of which is variable. HAD hydrolases are sometimes referred to as belonging to the DDDD superfamily of phosphohydrolases. Pssm-ID: 319768 [Multi-domain] Cd Length: 204 Bit Score: 254.39 E-value: 2.70e-87
|
||||||||
| Utr4 | COG4229 | Enolase-phosphatase E1 involved in merthionine salvage [Amino acid transport and metabolism]; |
2-167 | 1.21e-68 | ||||
Enolase-phosphatase E1 involved in merthionine salvage [Amino acid transport and metabolism]; Pssm-ID: 443373 [Multi-domain] Cd Length: 227 Bit Score: 207.70 E-value: 1.21e-68
|
||||||||
| enolase-ppase | TIGR01691 | 2,3-diketo-5-methylthio-1-phosphopentane phosphatase; This enzyme is the enolase-phosphatase ... |
13-167 | 7.53e-61 | ||||
2,3-diketo-5-methylthio-1-phosphopentane phosphatase; This enzyme is the enolase-phosphatase of methionine salvage, a pathway that regenerates methionine from methylthioadenosine (MTA). Adenosylmethionine (AdoMet) is a donor of different moieties for various processes, including methylation reactions. Use of AdoMet for spermidine biosynthesis, which leads to polyamine biosynthesis, leaves MTA as a by-product that must be cleared. In Bacillus subtilis and related species, this single protein is replaced by separate enzymes with enolase and phosphatase activities. [Central intermediary metabolism, Sulfur metabolism] Pssm-ID: 273760 [Multi-domain] Cd Length: 220 Bit Score: 187.74 E-value: 7.53e-61
|
||||||||
| HAD-SF-IA-v1 | TIGR01549 | haloacid dehalogenase superfamily, subfamily IA, variant 1 with third motif having Dx(3-4)D or ... |
15-135 | 2.04e-23 | ||||
haloacid dehalogenase superfamily, subfamily IA, variant 1 with third motif having Dx(3-4)D or Dx(3-4)E; This model represents part of one structural subfamily of the Haloacid Dehalogenase (HAD) superfamily of aspartate-nucleophile hydrolases. The superfamily is defined by the presence of three short catalytic motifs. The subfamilies are defined based on the location and the observed or predicted fold of a so-called "capping domain", or the absence of such a domain. Subfamily I consists of sequences in which the capping domain is found in between the first and second catalytic motifs. Subfamily II consists of sequences in which the capping domain is found between the second and third motifs. Subfamily III sequences have no capping domain in either of these positions.The Subfamily IA and IB capping domains are predicted by PSI-PRED to consist of an alpha helical bundle. Subfamily I encompasses such a wide region of sequence space (the sequences are highly divergent) that representing it with a single model is impossible, resulting in an overly broad description which allows in many unrelated sequences. Subfamily IA and IB are separated based on an aparrent phylogenetic bifurcation. Subfamily IA is still too broad to model, but cannot be further subdivided into large chunks based on phylogenetic trees. Of the three motifs defining the HAD superfamily, the third has three variant forms: (1) hhhhsDxxx(x)(D/E), (2) hhhhssxxx(x)D and (3) hhhhDDxxx(x)s where _s_ refers to a small amino acid and _h_ to a hydrophobic one. All three of these variants are found in subfamily IA. Individual models were made based on seeds exhibiting only one of the variants each. Variant 1 (this model) is found in the enzymes phosphoglycolate phosphatase (TIGR01449) and enolase-phosphatase. These three variant models (see also TIGR01493 and TIGR01509) were created withthe knowledge that there will be overlap among them - this is by design and serves the purpose of eliminating the overlap with models of more distantly relatedHAD subfamilies caused by an overly broad single model. [Unknown function, Enzymes of unknown specificity] Pssm-ID: 273686 [Multi-domain] Cd Length: 164 Bit Score: 90.53 E-value: 2.04e-23
|
||||||||
| HAD_L2-DEX | cd02588 | L-2-haloacid dehalogenase; L-2-Haloacid dehalogenase catalyzes the hydrolytic dehalogenation ... |
38-168 | 3.11e-06 | ||||
L-2-haloacid dehalogenase; L-2-Haloacid dehalogenase catalyzes the hydrolytic dehalogenation of L-2-haloacids to produce the corresponding D-2-hydroxyacids with an inversion of the C2-configuration. 2-haloacid dehalogenases are of interest for their potential to degrade recalcitrant halogenated environmental pollutants and their use in the synthesis of industrial chemicals. This family belongs to the haloacid dehalogenase-like (HAD) hydrolases, a large superfamily of diverse enzymes that catalyze carbon or phosphoryl group transfer reactions on a range of substrates, using an active site aspartate in nucleophilic catalysis. Members of this superfamily include 2-L-haloalkanoic acid dehalogenase, azetidine hydrolase, phosphonoacetaldehyde hydrolase, phosphoserine phosphatase, phosphomannomutase, P-type ATPases and many others. HAD hydrolases are found in all three kingdoms of life, and most genomes are predicted to contain multiple HAD-like proteins. Members possess a highly conserved alpha/beta core domain, and many also possess a small cap domain, the fold and function of which is variable. HAD hydrolases are sometimes referred to as belonging to the DDDD superfamily of phosphohydrolases. Pssm-ID: 319787 [Multi-domain] Cd Length: 216 Bit Score: 45.34 E-value: 3.11e-06
|
||||||||
| HAD_like | cd01427 | Haloacid dehalogenase-like hydrolases; The haloacid dehalogenase-like (HAD) superfamily ... |
47-142 | 6.13e-06 | ||||
Haloacid dehalogenase-like hydrolases; The haloacid dehalogenase-like (HAD) superfamily includes L-2-haloacid dehalogenase, epoxide hydrolase, phosphoserine phosphatase, phosphomannomutase, phosphoglycolate phosphatase, P-type ATPase, and many others. This superfamily includes a variety of enzymes that catalyze the cleavage of substrate C-Cl, P-C, and P-OP bonds via nucleophilic substitution pathways. All of which use a nucleophilic aspartate in their phosphoryl transfer reaction. They catalyze nucleophilic substitution reactions at phosphorus or carbon centers, using a conserved Asp carboxylate in covalent catalysis. All members possess a highly conserved alpha/beta core domain, and many also possess a small cap domain, the fold and function of which is variable. Members of this superfamily are sometimes referred to as belonging to the DDDD superfamily of phosphohydrolases. Pssm-ID: 319763 [Multi-domain] Cd Length: 106 Bit Score: 43.15 E-value: 6.13e-06
|
||||||||
| Blast search parameters | ||||
|
