Quality standards for assessment of iPSC in the field
This section is intended to provide a succinct overview of several assays for the quality control of human pluripotent stem cells and is neither comprehensive nor complete. No single example of these tests for pluripotency can be regarded as definitive on its own and we suggest that multiple methods can be used to integrate the various kinds of evidence into a comprehensive “life cycle” picture of each hPSC line ().
Life cycle of a pluripotent stem cell line. An hPSC line progresses through general stages throughout its life cycle: establishment, experimentation, long-term modification and long-term propagation. An hPSC line can “fork” into different (more...)
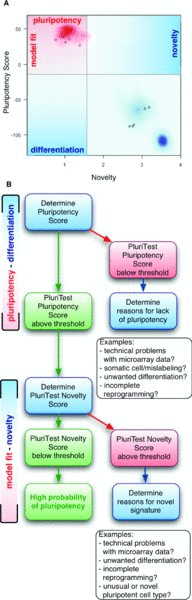
Our primary objective is to replace the teratoma assay, but PluriTest could replace other assays as well. We propose an alternative to the teratoma assay, which is expensive and requires animal surgery, using a purely data-driven standardized bioinformatic analysis of genome-wide transcriptional profiles. We provide background information, step-by-step instructions and exemplary interpretations for PluriTest as a prototypic example of such an assay.
The pluripotent stem cell field is expanding rapidly, and surrogate assays for assessing pluripotency, such as PluriTest, are critical for preclinical and translational stem cell research for the foreseeable future. However, proxy assays that rely on a defined set of biomarkers, such as RTPCR panels, immunocytochemistry markers, and methylation status of specific genes, lack the specificity required for selecting only truly pluripotent stem cells (Williams, Schuldt et al., 2011). For example, a positive result using immunocytochemistry or RTPCR for pluripotency-associated genes indicates that pluripotent cells are present, but provides no information about the homogeneity of the population; even 14-day old embryoid bodies are identified as pluripotent by these assays. Similarly, assessment of the methylation status of the promoter of a pluripotency-associated gene would indicate pluripotency for some cancer cells, partially reprogrammed cells, and karyotypically abnormal cells. PluriTest avoids these pitfalls by comparing the whole genome expression profile of the query sample with a model of confirmed undifferentiated euploid hPSCs built from a very large dataset gene expression information (Williams, Schuldt et al., 2011). This assay not only identifies samples as containing pluripotent cells, but also reveals contamination by differentiated cells, and genomic and epigenomic abnormalities that affect gene expression patterns.
PluriTest is a tool for rapid assessment of pluripotency in human in vitro stem cell preparations. We expect that researchers will initially screen for promising stem cell colonies after derivation or experimental manipulation by using RTPCR or immunocytochemistry for detecting pluripotency-associated markers before confirming pluripotency with whole genome expression analysis for PluriTest (). However, with near-perfect sensitivity and specificity, PluriTest can function as the sole means to determine pluripotency in any given culture. We envision that further developments of PluriTest platform will have utility for not only testing for pluripotency, but also identifying differentiation propensity and genomic integrity in stem cell preparations.
Cell culture morphology and dynamics
hESCs and hiPSCs show a distinct colony morphology when cultured on murine embryonic fibroblasts (Thomson, Itskovitz-Eldor et al., 1998; Takahashi, Tanabe et al., 2007). This characteristic feature allows for the identification of successfully reprogrammed iPSCs (Meissner, Wernig et al., 2007) ().
Basic quality control of iPSC colonies. Selecting “good” colonies from “bad” ones is probably the most commonly used quality control step in hPSC research. Here we show iPSCs generated from fibroblasts with episomal vectors (more...)
Visual inspection and manual selection of “good” from “ugly” looking colonies probably remains the most under-appreciated yet most important control instrument for pluripotent quality assessment, used every day in hPSC labs worldwide. Development of the expertise to decide which colonies to pick and which to discard requires apprenticeship with experienced researchers and is difficult to operationalize (Loring, Schwartz et al., 2007). Typical hPSC morphological characteristics are often lost or altered in modern, automatable feeder-free and defined culture conditions (Ludwig, Bergendahl et al., 2006; Wang, Schulz et al., 2007); this is one of the reasons that most preclinical research labs routinely culture and derive hPSCs on feeder cell systems.
Karyotypic and genomic stability
hPSCs need to be karyotyped on a regular basis (Loring, Schwartz et al., 2007). Mounting evidence shows that PSC regularly acquire a pluripotency-specific set of karyotypic abnormalities (Mayshar, Ben-David et al., 2010; Laurent, Ulitsky et al., 2011). Unfortunately, subchromosomal mutations frequently occur in the same genomic regions that are found to be amplified on the karyotypic level (Laurent, Ulitsky et al., 2011). Such small abnormalities can only be detected with genome wide, high resolution SNP (single nucleotide polymorphism) and CGH (comparative genome hybridization) microarrays, or DNA sequencing. Recent evidence suggests that when compared to their respective parental cells, some iPSC lines contain 1000 to 2000 single allele base changes, including more than 10 non-synonymous changes in coding genes(Cheng, Hansen et al., 2012). These data highlight the dynamic character inherent to the life cycle of a hPSC line.
Marker expression (RT-PCR, IHC, cell surface markers)
Almost every manuscript that reports the derivation of hESC and iPSC lines shows immunocytochemical proof of expression of pluripotency-associated markers such as POU5F1 (OCT4), SOX2, NANOG, and/or the antigens TRA1-61, TRA1-81, and SSEA4. Several companies offer “stem cell characterization kits” with these antibody panels. Other commercial assays are based on expression measurements for a group of pluripotency-associated genes. The International Stem Cell Initiative proposed a set of 96 QRT-PCR markers for the assessment of pluripotency and differentiation in hESC lines (Adewumi, Aflatoonian et al., 2007). Commercialized versions of these stem cell biomarker sets are able to positively identify cells as hPSCs, but cannot reliably discriminate aberrant phenotypes such as partially reprogrammed iPSCs, epigenetically or genetically abnormal iPSCs and hESCs, or teratocarcinoma cell lines.
There is a recent trend toward using cell surface markers, such as antibodies (Lowry, Richter et al., 2008; Chan, Ratanasirintrawoot et al., 2009) and lectins (Wang, Nakagawa et al., 2011) that selectively bind to hESCs and iPSCs. The advantage of these markers is that they can be used to sort cells by flow cytometry or magnetic bead-based technology. The magnetic bead technology may become an important technological development to support fully automated high throughput hiPSC derivation and culture methods, and it has already been integrated into improved protocols for automated hiPSC selection (Dick, Matsa et al., 2011; Valamehr, Abujarour et al., 2012) and hPSC passaging.
In vitro differentiation
Numerous in vitro differentiation protocols for all three germ layer lineages and many terminally differentiated somatic cell types have been reported. However, there is no current standard of a single set of in vitro differentiation protocols necessary or sufficient to demonstrate pluripotent differentiation potential. Undirected differentiation into all three germ layers is often performed using an embryoid body protocol (Martin and Evans et al., 1975), in which the hPSCs are aggregated and cultured in suspension until the cells begin to differentiate. There is no common protocol for generating EBs, and no commonly agreed-upon set of markers for quantitatively analyzing the outcome of such an experiment. Even with the comprehensive ISCI 96 stem cell marker set, it has been difficult to reliably discern EBs from undifferentiated hESC cultures, possibly due to residual undifferentiated PSC even in late EB stages (Adewumi, Aflatoonian et al., 2007). Attempts have been made to standardize EB formation (Burridge, Anderson et al., 2007; Pick, Azzola et al., 2007; Ungrin, Joshi et al., 2008), but no commonly accepted standard has emerged so far.
In vivo differentiation (teratoma assay)
The teratoma assay for in vivo demonstration of pluripotential differentiation capabilities based on work by Stevens and Little in the 1950s and 1960s (Stevens and Little et al., 1954; Stevens et al., 1964) was employed in the first-of-its-kind manuscripts demonstrating the derivation of murine and human ESC lines (Evans and Kaufman et al., 1981; Martin et al., 1981; Thomson, Itskovitz-Eldor et al., 1998) as well as murine and human iPSC lines (Takahashi and Yamanaka et al., 2006; Takahashi, Tanabe et al., 2007; Yu, Vodyanik et al., 2007). The assay requires the sacrifice of laboratory animals and is expensive and time consuming (Peterson, Tran et al., 2011).
Although the teratoma assay is regarded by some researchers as still essential for the qualification of new human pluripotent cell lines (Dolgin et al., 2010), in practice it is actually performed for less than half of the new hESC or hiPSC lines reported. In a recent study, we found that from 1998 to 2009 only 44% of 639 hESC lines and only 36% of 777 iPSC lines were actually tested in vivo by the teratoma assay (Müller, Goldmann et al., 2010).
For producing teratomas, usually about on million hPSC are in injected into each of three immunodeficient male mice. Common injection sites are under the skin, into the hind limb muscles, into the kidney capsule, into the testes; one group transplants cells to the spinal cord (Israel, Yuan et al., 2012). The animals need to be monitored on a daily basis for two to three months and their greatest potential for suffering is at the end of that time period, when teratomas have significantly increased in size often due to fluid-filled cysts, with potential for compression of internal organs (Peterson, Tran et al., 2011) and sometimes resulting in ulceration in tumors formed by subcutaneous and intramuscular injection.
The analysis of teratomas is not straightforward and requires the expertise of a trained pathologist (Gertow, Cedervall et al., 2011). Often, the most benign type of teratomas (called mature or cystic) turns out the least informative. Growth of large fluid-filled cysts suppresses the development of the surrounding stem cell-derived tissues, due to pressure atrophy, resulting in teratomas without discernible differentiation of some germ layer derivatives. Analysis of cellular differentiation in teratomas is usually superficial and qualitative, with most researchers only highlighting islands of identifiable differentiation based on histological features, such as clusters of neural rosettes or secretory glandular tissue (Müller, Goldmann et al., 2010). In our experience, 30 to 80% of the observable teratoma tissue cannot be confidently assigned to a specific germ layer with conventional histological methods .
Most mature lineages cannot be confidently modeled and observed within teratomas. For example, neural derivatives usually do not progress beyond an immature embryonic or fetal state. Consequently, for disease modeling applications or drug screening efforts requiring mature differentiated cell products such as spinal cord motoneurons (Dimos, Rodolfa et al., 2008) or differentiated cells from body surfaces such as epidermal keratinocytes, the teratoma assay has no informative value for the propensity of an iPSC line to differentiate into these cell types.
Genome wide profiling
Genome wide profiles of pluripotent stem cells were published for the first time in 2001 (Loring, Porter et al., 2001). Since then, profiling of stem cell transcriptomes has become a commonly used methodology for the analysis of newly established hPSC lines to compare them with existing hESC or iPSC lines. In regard to global profiling methods, we propose the distinction of functional versus structural assays. Transcriptomics currently is probably the prime example for a genome wide functional assay. As a highly integrated molecular machinery in each cell is necessary for transcribing snippets of DNA in a genomic locus into various kinds of RNA (mRNA, microRNA and nclRNA), transcriptional profiles are the consequence of an active and highly orchestrated process within each cell. In contrast, epigenomic profiles are not well understood and the data generated by genome wide profiling, of CpG marks for example, cannot yet be directly linked with a relevant cellular function. A gene that has been found to be over-expressed in one hiPSC line and hypothetically connected to a specific function can be easily tested further through over-expression or knockdown of the same gene in several hPSC lines. The same is currently impossible for differentially methylated regions found in one hPSC line when compared to others. Eventually, epigenetic signals will surely be linked with processes governed by and governing cellular states, but currently our knowledge of the epigenetic makeup of stem and somatic cells assays can be interpreted only as structural variations without immediate and obvious insights or easily testable hypotheses.
While it is in principle possible to establish pluripotency assays with all types of genome-wide data such as mirRNA (Laurent, Chen et al., 2008), mRNA (Müller, Laurent et al., 2008; Müller, Schuldt et al., 2011), chromatin modifications (Ernst, Kheradpour et al., 2011) and CpG methylation (Laurent, Wong et al., 2010; Bock, Kiskinis et al., 2011; Nazor, Altun et al., 2012 (in press)), the only currently biologically interpretable signatures are those based on mRNA transcriptomes.